
肉质扁脑珊瑚的有性繁殖及早期发育
作者简介:肖宝华(1978—), 男, 福建省长乐市人, 硕士, 主要从事海洋生态学研究。电话: 0759-2396216, E-mail:gdouxxhpaper@126.com
收稿日期: 2016-04-28
要求修回日期: 2016-09-06
网络出版日期: 2017-01-19
基金资助
广东省公益研究与能力建设专项 (K15216)
广东省海洋渔业科技推广专项(A201308E02)
大鹏新区产业发展专项(DPKJ201500080)
Sexual reproduction and early development of Platygyra carnosus
Received date: 2016-04-28
Request revised date: 2016-09-06
Online published: 2017-01-19
Supported by
Public Welfare Research and Capacity Building of Guangdong Province (K15216)
Marine Fishery Science and Technology Promotion Project in Guangdong Province (A201308E02)
Dapeng New District Development Projects (DPKJ201500080)
Copyright
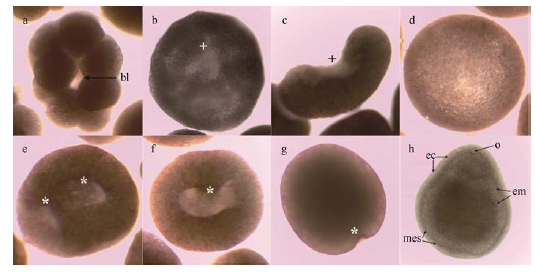
在实验室养殖的条件下对肉质扁脑珊瑚Platygyra carnosus的有性繁殖进行初步观察, 收集其排放的精卵进行受精, 同时记录了肉质扁脑珊瑚早期发育各个时期的形态特征。观察结果表明, 肉质扁脑珊瑚为雌雄同体, 体外受精, 在月圆后第9天开始出现排卵现象, 第11天排卵量达到最大。肉质扁脑珊瑚的卵为粉红色圆球形。在水温27±0.5℃的条件下, 肉质扁脑珊瑚早期发育共经历5个发育阶段17个时期, 受精卵经48h发育为浮浪幼虫, 96h以后开始寻找基底进行附着。本研究为国内首次记录和报道肉质扁脑珊瑚的有性繁殖及胚胎发育过程, 可以为石珊瑚的生态修复积累理论依据。
肖宝华 , 廖宝林 , 杨小东 , 谢子强 . 肉质扁脑珊瑚的有性繁殖及早期发育[J]. 热带海洋学报, 2017 , 36(1) : 65 -71 . DOI: 10.11978/2016044
Sexual reproduction of Platygyra carnosus was observed preliminarily under the condition of artificial culture. Eggs and sperms of Platygyra carnosus were collected when they were spawned. The morphological characteristics in early development of Platygyra carnosus during different periods were recorded and described. The results showed that the corals were hermaphrodites, in vitro fertilization, and began spawning on the ninth day after the full moon,, then the spawning quantity of corals increased to the maximum on the 11th day after the full moon. The eggs of Platygyra carnosus were spherical in shape and pink in color. The early development process underwent five phases and 17 stages under the conditions of 27±0.5℃. The fertilized eggs developed into planula in 48 h and the larvae began to settle after 96 h. The sexual reproduction and early development of Platygyra carnosus were reported for the first time in China.
Key words: Platygyra carnosus; sexual reproduction; early development
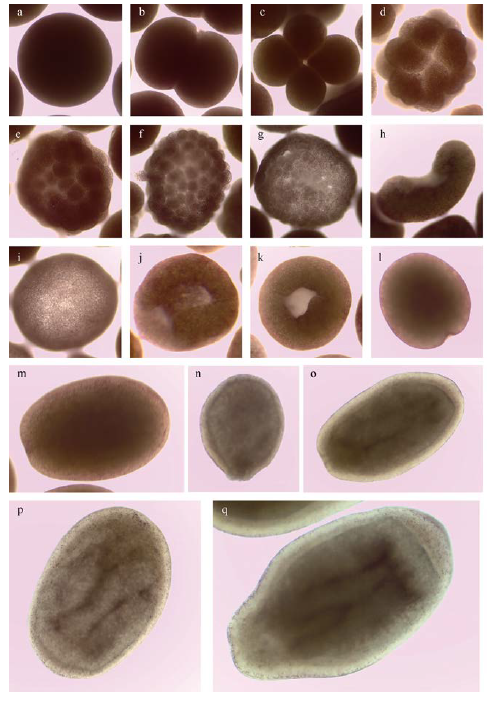
Tab. 1 Early development of Platygyra carnosus表1 肉质扁脑珊瑚早期发育过程 |
| 受精后时间 | 胚胎发育时期 | 主要发育特征 | 图示 |
|---|---|---|---|
| 0 | 受精卵 | 受精卵呈粉红色圆球状 | |
| 2h | 2细胞期 | 受精卵分裂成大小相等的2个细胞 | |
| 3h30min | 4细胞期 | 受精卵进一步分裂成4个细胞 | |
| 4h30min | 16细胞期 | 受精卵进一步分裂成16个细胞, 分裂球在胚胎的分布变得不规则 | |
| 5h30min | 32细胞期 | 受精卵进一步分裂成32个细胞 | |
| 6h | 桑葚胚期 | 受精卵进一步分裂成64~128个细胞,细胞团与桑葚球非常相似, 胚胎呈不规则形态 | |
| 7h | 囊胚早期 | 受精卵进一步分裂成128个以上的细胞, 此时胚胎表面变得平滑, 中央出现细胞内陷, 形成假胚孔 | |
| 8h | 囊胚中期 | 囊胚进一步分裂, 胚胎变得扁平, 发育为坐垫状, 凹面继续内陷, 假胚孔加深且表面更加平滑 | |
| 9h | 囊胚晚期 | 坐垫状囊胚逐渐收拢形成一个球状胚体, 假胚伴随此过程逐渐孔消失 | |
| 15h | 原肠胚早期 | 在极短时间内胚胎表面形成两个胚孔 | |
| 17h | 原肠胚中期 | 两个胚孔进一步扩大并最终融合成一个胚孔, 胚孔内陷形成原口, 由于细胞的迁移开始形成两个胚层的分化 | |
| 24h | 原肠胚晚期 | 原肠胚的原口开始逐渐闭合, 胚体逐渐拉长成椭圆形或梨形 | |
| 36h | 原肠胚末期 | 原肠作用临近结束, 原口封闭完成, 胚胎发育结束 | |
| 48h | 浮浪幼虫早期 | 浮浪幼虫维持椭圆形或梨形的形态, 此时外胚层已发育出鞭毛的结构, 已经具备游泳能力 | |
| 58h | 浮浪幼虫中期 | 浮浪幼虫形态明显, 形态拉长为圆柱状, 可在显微镜底下清晰观察到胚体内两个胚层, 幼虫开始四处游动 | |
| 84h | 浮浪幼晚期 | 浮浪幼虫内部分化明显, 内胚层发育出肠系膜 | |
| 96h | 浮浪幼末期 | 游动能力加强, 开始尝试寻找基质附着 |
Fig. 1 Morphology of early development of P. carnosus |
Fig. 2 Morphology of early development of P. carnosus. |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}