
三亚湾近岸浮游纤毛虫群落的昼夜变化特征*
作者简介:王有军(1990—), 男, 山东省日照市人, 硕士研究生, 从事浮游动物生态学研究。E-mail:wangyoujun13@mails.ucas.ac.cn
收稿日期: 2016-04-05
要求修回日期: 2016-05-13
网络出版日期: 2017-01-19
基金资助
中国科学院战略性先导科技专项子课题(XDA13020102)
国家自然科学基金项目(41276160)
广东省科技计划项目(2015A020216013)
Diel variation of community structure of planktonic ciliates in coastal ecosystem of Sanya Bay
Received date: 2016-04-05
Request revised date: 2016-05-13
Online published: 2017-01-19
Supported by
Strategic Priority Research Programof the Chinese Academy of Sciences (XDA13020102)
National Natural Science Foundation of China (41276160)
Science and Technology plan Projects of Guangdong Province (2015A020216013)
Copyright
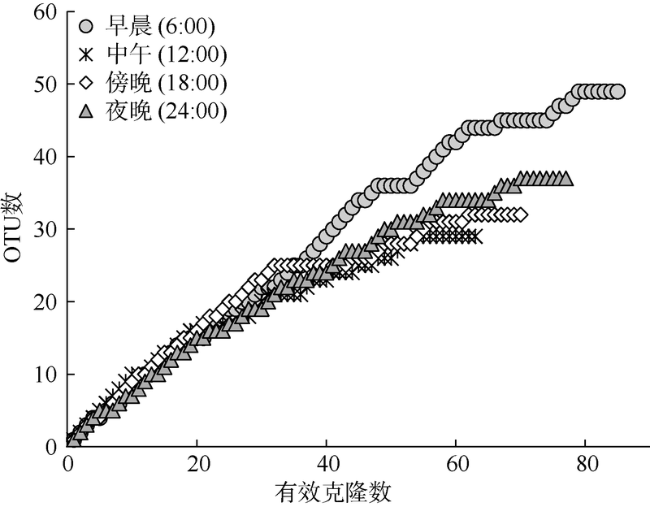
浮游纤毛虫广泛分布于海洋环境中, 是连接微食物环和经典食物链的关键环节, 在生态系统的物质传递和能量流通中占据重要地位。为深入认识纤毛虫在热带海区的分布特征及其生态功能, 于2014年4月在三亚湾近岸海区设置昼夜连续监测站点, 分早晨(6:00)、中午(12:00)、傍晚(18:00)和夜晚(24:00)四个时间点分别采集浮游纤毛虫及环境因子检测样品, 并用分子生物学方法分析了表层水体浮游纤毛虫群落结构的昼夜变化特征。结果显示: 1)表层浮游纤毛虫多样性高, 共获得79条不同代表序列; 2)不同时间表层海水中浮游纤毛虫多样性差异显著, 表现为白天降低, 夜晚升高, 其多样性高低依次为: 早晨 [48个操作分类单元(operational taxonomic unit, OTU)] >夜晚 (37个OTU) > 傍晚 (32个OTU) > 中午 (29个OTU)。浮游纤毛虫群落结构表现出昼夜变化现象, 可能是因为不同类群纤毛虫具有不同的空间分布规律。
王有军 , 徐翠莲 , 胡思敏 , 李涛 , 刘胜 . 三亚湾近岸浮游纤毛虫群落的昼夜变化特征*[J]. 热带海洋学报, 2017 , 36(1) : 72 -80 . DOI: 10.11978/2016034
Planktonic ciliates are widespread micro-zooplankton in marine ecosystem and are considered as a key link between microbial loop and classic food chain as they play important roles in transfer processes of material and energy in the ecosystem. To better understand the distribution and ecological function of planktonic ciliates in tropical ecosystem, ciliate samples were collected in the coastal waters of Sanya Bay at 6:00, 12:00, 18:00 and 24:00, respectively, in April 2014. Species diversity and community structure of ciliate from different times were analyzed by molecular detection based on species-specific primers. The results were as follows. 1) Planktonic ciliate showed high diversity, with about 79 operational taxonomic units (OTUs) detected. 2) Significant variation of species diversity was uncovered at different times during a day, showing a decrease trend in day time but increase trend at night time and the order was morning [48 OUTs(operational taxonomic units)] > night (37 OTUs) > evening (32 OTUs) >midday (29 OTUs). The reason may be that different ciliate populations have different distribution modes.
Key words: Sanya Bay; planktonic ciliates; diversity; diel variation
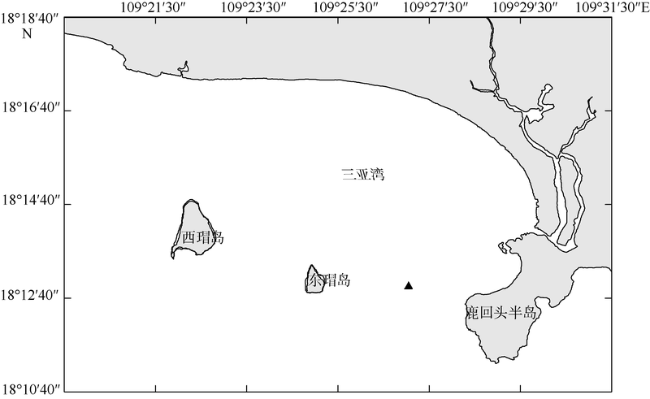
Fig. 1 Sampling location in Sanya Bay图1 三亚湾调查站位(▲) |
Tab. 1 Diel variation of environmental factors at the sampling site of Sanya Bay表1 三亚湾近岸站位环境因子昼夜变化 |
| Chl a含量/(μg∙L-1) | POM/(mg∙L-1) | |
|---|---|---|
| 早晨(6:00) | 0.76 | 1.05 |
| 中午(12:00) | 0.80 | 0.63 |
| 傍晚(18:00) | 2.09 | 1.10 |
| 夜晚(24:00) | 6.69 | 1.43 |
注: “Chla” 表示叶绿素a; “POM” 表示颗粒有机物。 |
Tab. 2 Diversity indices of planktonic ciliate at different times based on the samples analyzed表2 不同时间点样品的测序状况 |
| 送测克隆数 | 有效克隆数 | 辛普森指数 | 香农-威纳指数 | |
|---|---|---|---|---|
| 早晨(6:00) | 90 | 85 | 0.9686 | 3.685 |
| 中午(12:00) | 70 | 63 | 0.9393 | 3.073 |
| 傍晚(18:00) | 80 | 70 | 0.9359 | 3.107 |
| 夜晚(24:00) | 80 | 77 | 0.9489 | 3.325 |
| 合计 | 320 | 295 | --- | --- |
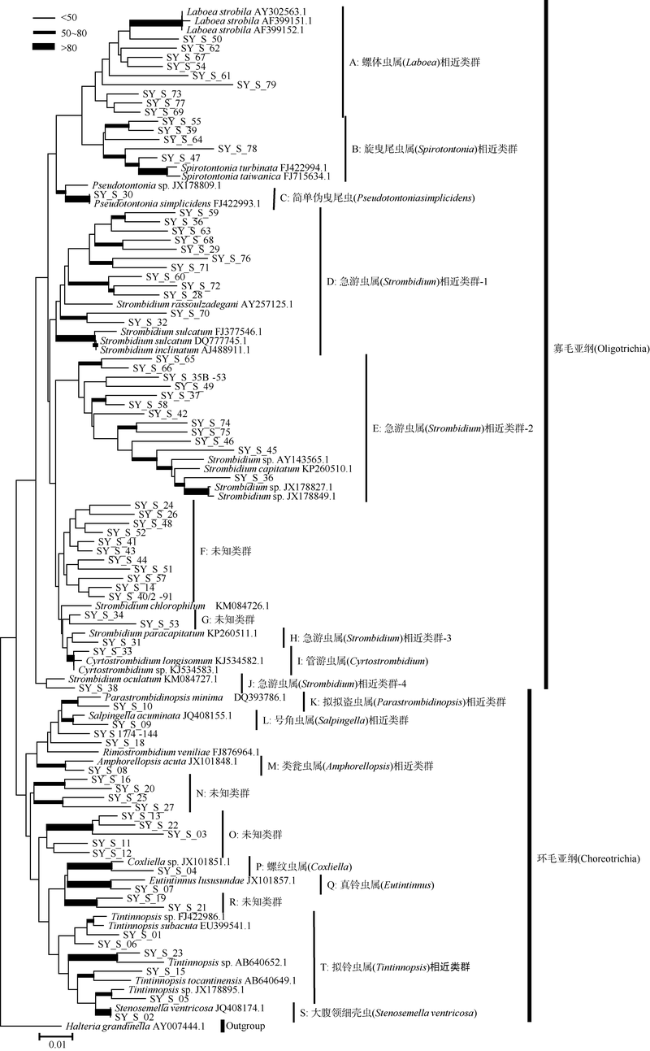
Fig. 2 Neighbor joining tree of 18S rDNA sequences retrieved from the ciliate samples. Shown at nodes are bootstrap confidence values based on 1000 replicates图2 基于从现场纤毛虫多样性样品中获得的序列和GenBank中匹配到的相似序列比对后构建的NJ进化树 |
Fig. 3 Diel variation of ciliate species composition at the study site (S) in Sanya Bay图3 三亚湾近岸S站位不同时间浮游纤毛虫的群落组成的昼夜变化 |
Fig. 4 Rarefaction curves to indicate diversity coverage of our sequencing scales for different clone libraries图4 4个时间点送测克隆数所覆盖的浮游纤毛虫种类 |
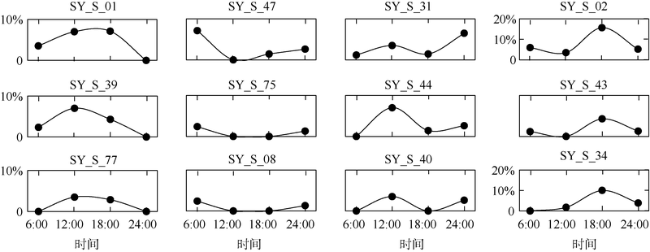
Fig. 5 Diel distribution changes of different ciliates in the surface layers of Sanya Bay图5 三亚湾S站位表层不同浮游纤毛虫的昼夜分布规律 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China(AQSIQ), 2008. GB/T 12763.6-2007 Specifications for oceanographic survey—Part 6:Marine biological survey[S]. Beijing: China Standard Publishing House (in Chinese).
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}