
长期高氨氮暴露对海水青鳉毒理效应的蛋白质组学研究
作者简介:高娜(1988—), 女, 山东省新泰市人, 博士研究生, 主要从事海洋生物学研究。E-mail: gaona12@mails.ucas.ac.cn
收稿日期: 2016-12-02
要求修回日期: 2017-04-21
网络出版日期: 2017-09-22
基金资助
国家重点基础研究发展规划项目(2015CB452904)
国家自然科学基金项目(41376161、31501862)
中国科学院百人计划项目
广东省科技计划项目(2014B030301064)
Proteomic responses of marine medaka (Oryzias melastigma) to chronic high environment ammonia exposure
Received date: 2016-12-02
Request revised date: 2017-04-21
Online published: 2017-09-22
Supported by
State Key Development Program for Basic Research of China (2015CB452904)
National Natural Science Foundation of China (41376161, 31501862)
100 Talents Program of the Chinese Academy of Sciences to ZHANG Li
Science and Technology Planning Project of Guangdong Province, China (2014B030301064).
Copyright
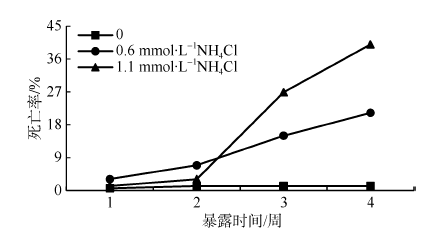
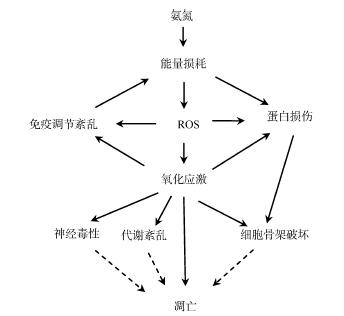
氨氮是大多数硬骨鱼的氮代谢产物, 但是环境中氨氮浓度过高也会对鱼类产生毒性作用。该研究结果显示, 海水青鳉暴露于0.6和1.1mmol·L-1 NH4Cl 4周后的死亡率分别为21.25%和40%。这说明高浓度氨氮暴露对海水青鳉具有明显的毒性作用, 而且暴露浓度越高, 毒性越大。利用蛋白质组学的研究技术研究海水青鳉在长期氨氮暴露后的蛋白质响应变化, 发现在0.6mmol·L-1 NH4Cl暴露条件下有16种蛋白的表达发生明显的变化, 而在1.1mmol·L-1 NH4Cl暴露条件下有7种蛋白的表达发生明显的变化。这些蛋白主要参与氧化应激的产生, 影响海水青鳉细胞结构组成、肌肉收缩、视觉通路、代谢调节和免疫调节, 引起神经毒性。该研究利用蛋白质组学的研究方法研究了长期氨氮暴露对海水青鳉的毒理效应, 为利用蛋白质组学探讨环境污染物的毒性效应提供了更深刻的认识。
高娜 , 朱丽梅 , 于德良 , 张黎 . 长期高氨氮暴露对海水青鳉毒理效应的蛋白质组学研究[J]. 热带海洋学报, 2017 , 36(5) : 40 -48 . DOI: 10.11978/2016126
Abstract: Most teleost fish excretes nitrogen waste as ammonia, but high environment ammonia is toxic to fish. In this study, we demonstrated that the death rate of marine medaka Oryzias melastigma exposed to 0.6 and 1.1 mmol·L-1 NH4Cl for four weeks were 21.25% and 40.00%, respectively, suggesting high environment ammonia is toxic to the fish. Besides, the higher concentration the ambient ammonia, the more toxic it is to the fish. We investigated the responses of chronic ammonia exposure in marine medaka using proteomics. Sixteen proteins were found to be remarkably altered in the fish exposed to 0.6 mmol·L-1 NH4Cl and seven proteins, in the fish exposed to 1.1 mmol·L-1 NH4Cl. The function of these proteins included inducing oxidative stress, neurotoxicity, disturbance in cell structure, muscle contraction, visual pathway, metabolic and immunological regulation. This is the first report of studying the toxicological effect of ammonia on marine medaka using proteomics. It provides important insights into toxicological effects of environmental contaminant using proteomics.
Key words: chronic ammonia exposure; proteomic; marine medaka; toxicological effect
Fig. 1 Death rate of marine medaka exposed to 0, 0.6 and 1.1mmol·L-1 NH4Cl during 4-week exposure图 1 暴露于0、0.6 和 1.1mmol·L-1 NH4Cl 的海水青鳉在4周期间的死亡率 |
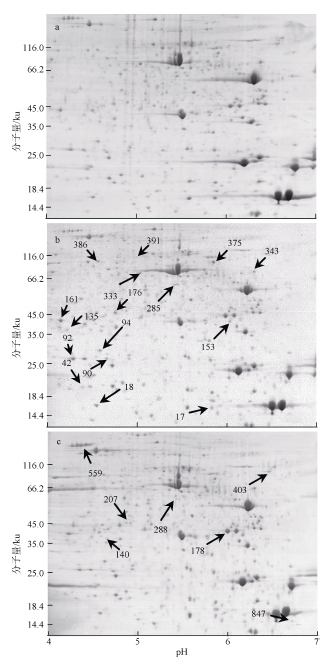
Fig. 2 Representive 2-DE images of proteins extracted from marine medaka in control group (a), exposed to 0.6mmol·L-1 NH4Cl (b) and exposed to 1.1mmol·L-1 NH4Cl (c)图2 氨氮暴露后海水青鳉双向电泳图谱 |
Tab. 1 List of proteins which were differentially expressed in O. melastigma induced by chronic ammonia exposure表1 长期氨氮暴露后海水青鳉体内差异表达蛋白质谱鉴定结果 |
| 蛋白 点数 | 组别 | 蛋白名称 | 物种 | 序列号 | 分子量/u | 等电点 | 蛋白 得分 | 序列 覆盖率 | 匹配 肽数 | 倍数 变化 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 氧化 应激 | 94 | A | 低分子量蛋白: 类线粒体的 过氧化物还原酶5 | 青鳉 | gi|432899770 | 20491 | 8.68 | 114 | 0.18 | 2 | 4.26 | |
| 140 | B | 类蛋白酶体β亚基4 | 青鳉 | gi|432881057 | 28758 | 6.32 | 138 | 0.20 | 3 | 2.58 | ||
| 178 | B | 胰蛋白酶原 | 青鳉 | gi|145966014 | 26708 | 5.16 | 255 | 0.12 | 1 | 1.63 | ||
| 细胞结构 | 285 | A | 类透明质酸蛋白聚糖连接蛋白1 | 青鳉 | gi|432874420 | 37805 | 8.18 | 63 | 0.03 | 1 | -1.54 | |
| 288 | B | 类透明质酸蛋白聚糖连接蛋白1 | 青鳉 | gi|432874420 | 37805 | 8.18 | 76 | 0.03 | 1 | -1.71 | ||
| 肌肉收缩 | 92 | A | 类丝切蛋白-2 | 青鳉 | gi|432936587 | 18889 | 6.62 | 426 | 0.37 | 4 | 1.60 | |
| 343 | A | 类原肌球蛋白α1链亚型4 | 青鳉 | gi|432861692 | 31829 | 4.77 | 268 | 0.19 | 3 | -1.70 | ||
| 333 | A | 肌钙蛋白T, 类快骨骼肌型亚型2 | 青鳉 | gi|432860165 | 27699 | 9.41 | 96 | 0.10 | 2 | -1.55 | ||
| 403 | B | 类钙腔蛋白B | 青鳉 | gi|432863493 | 29610 | 4.42 | 152 | 0.18 | 3 | -1.92 | ||
| 视觉通路 | 176 | A | 中间丝蛋白ON3亚型2 | 青鳉 | gi|432864505 | 57350 | 5.06 | 69 | 0.02 | 1 | 1.63 | |
| 90 | A | α-晶状体A链 | 青鳉 | gi|432958610 | 19919 | 5.95 | 184 | 0.44 | 6 | 1.70 | ||
| 135 | A | 类β-晶状体B1链 | 青鳉 | gi|432878256 | 28885 | 8.36 | 70 | 0.23 | 4 | 4.88 | ||
| 161 | A | 类β-晶状体A2链亚型1 | 青鳉 | gi|432964694 | 23855 | 6.09 | 377 | 0.51 | 5 | 2.07 | ||
| 代谢调节 | 17 | A | 14ku载脂蛋白部分片段 | 海水青鳉 | gi|328963182 | 11405 | 6.23 | 134 | 0.32 | 1 | -1.53 | |
| 375 | A | 类线粒体ATP合成酶β亚基 | 青鳉 | gi|432849647 | 55238 | 5.10 | 562 | 0.24 | 6 | 2.15 | ||
| 386 | A | 类α-烯醇酶 | 青鳉 | gi|432866138 | 45323 | 6.36 | 431 | 0.30 | 5 | 1.99 | ||
| 207 | B | 类异戊烯基二磷酸Δ异构酶1 | 青鳉 | gi|432927313 | 32545 | 6.06 | 110 | 0.23 | 3 | -3.27 | ||
| 免疫调节 | 153 | A | 类组织蛋白酶B | 青鳉 | gi|432852559 | 37145 | 5.93 | 90 | 0.05 | 1 | 2.07 | |
| 42 | A | 类半乳凝素9 | 青鳉 | gi|432852360 | 23618 | 8.81 | 90 | 0.16 | 2 | 2.81 | ||
| 559 | B | 类胎球蛋白B | 大黄鱼 | gi|734621215 | 57367 | 5.99 | 63 | 0.01 | 1 | 2.75 | ||
| 神经毒性 | 18 | A | 类S100蛋白A | 青鳉 | gi|432908621 | 11360 | 5.62 | 60 | 0.08 | 1 | 1.90 | |
| 391 | A | 突触囊泡膜蛋白VAT-1同系物 | 青鳉 | gi|432922308 | 46708 | 5.40 | 150 | 0.10 | 3 | 1.53 | ||
| 847 | B | 类S100B亚型1 | 青鳉 | gi|432848584 | 11052 | 4.43 | 67 | 0.17 | 1 | -2.70 | ||
注: 组A: 0.6 mmol·L-1 NH4Cl 暴露组; 组B: 1.1 mmol·L-1NH4Cl 暴露组。 |
Fig. 3 The hypothetical model of the toxicity mechanism in the marine medaka to chronic ammonia exposure图3 长期氨氮暴露对海水青鳉产毒性效应的机制假设模型 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}