
光裸方格星虫增养殖对滩涂底质的影响
作者简介:李俊伟(1982—), 男, 河北省故城人, 助理研究员, 博士, 从事养殖生态学研究。E-mail: lijunwei303@163.com
收稿日期: 2017-04-05
要求修回日期: 2017-06-22
网络出版日期: 2018-02-02
基金资助
国家自然科学青年基金(41606137)
广东省自然科学基金(2015A030310260、2015A030313855)
广东省海洋渔业科技推广项目(B201601-01)
中央级公益性科研院所基本科研业务费专项资金(中国水产科学研究院南海水产研究所)项目(2013ZD01)
广东省渔业生态环境重点实验室开放基金(LFE-2016-9)
Influence of aquaculture of Sipunculus nudus on sediment
Received date: 2017-04-05
Request revised date: 2017-06-22
Online published: 2018-02-02
Supported by
National Natural Science Foundation of China (41606137)
Natural Science Foundation of Guangdong Province (2015A030310260, 2015A030313855)
Marine Fishery Science and Technology Industry Projects from Guangdong Province (B201601-01)
Scientific Research Funds for Central Non-profit Institutes, South China Sea Fisheries Research Institute (2013ZD01)
Guangdong Provincial Key laboratory of Fishery Ecology and Environment (LFE-2016-9)
Copyright
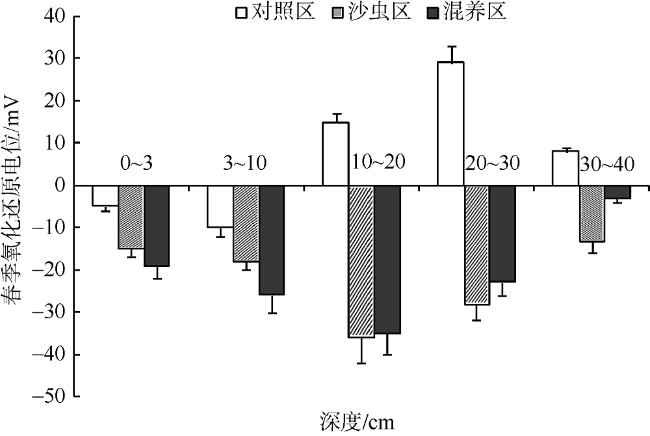
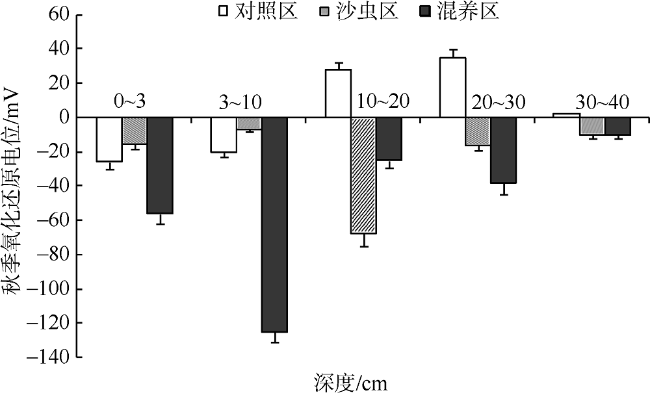
基于2016年春、秋两个季节的现场实验数据, 研究光裸方格星虫(Sipunculus nudus Linnaeus)滩涂增养殖对滩涂底质的影响。实验对方格星虫单养区(S组)、方格星虫-贝类混养区(P组)和无星虫对照区(C组)进行了有机质、粒径组成、氧化还原电位(ORP)等指标测定, 结果表明: 1) 秋季S组20~30cm层次有机质含量和细沙比例均显著高于春季, 而C组仅表层物质有所增加, 表明方格星虫具有较强的表层物质迁移能力, 且其迁移深度为20~30cm。2) 秋季C组各层氧化还原电位值(ORP)均无明显降低趋势, 而S组仅10~20cm层次有一定程度降低, 表明方格星虫增养殖并不会明显降低滩涂底质状况。3) P组各层有机质以及细沙含量增加, 表明皱肋文蛤的过滤沉积作用可以提高滩涂颗粒物质的积累, 且方格星虫混养会加速颗粒物质的向下迁移; P组皱肋文蛤的栖息层次(0~3cm、3~10cm) ORP明显低于S组(p<0.05), 表明高密度的皱肋文蛤代谢活动会加剧0~10cm的底质恶化。4) 方格星虫与皱肋文蛤具有生态互补作用, 在同一区域进行增养殖可以提高滩涂物质利用率及碳汇功能; 方格星虫扰动引入富氧水可以缓解皱肋文蛤栖息层次的底质。
李俊伟 , 颉晓勇 , 郭永坚 , 朱长波 , 陈素文 , 丁贤 , 李婷 , 陈振国 . 光裸方格星虫增养殖对滩涂底质的影响[J]. 热带海洋学报, 2018 , 37(1) : 37 -44 . DOI: 10.11978/2017037
To study the effects of culturing and proliferation of Sipunculus nudus on tidal flat sediment, we carried out an investigation in a culturing zone located in Suixi, Beibu Gulf, China in spring and autumn 2016. The organic matter content (OM), grain size and oxidation reduction potential (ORP) of the sediment were determined in the S group (Sipunculus nudus monoculture), P group (polyculture of S. nudus and Meretrix lyrata) and C group (blank zone without S. nudus). The OM and fine sand of the layer (20~30 cm) in the S group accumulated obviously, but only the surface layer (0~3 cm) in the C group had a similar tendency, which indicated that S. nudus can migrate down the surface sediment, and the migration depth was about 20~30 cm in the tidal flat. The ORP of the pore water in the C group had no significant decrease, and most of the layers in the S group had similar changing trends except that the layer of 10~20 cm decreased to some extent, which indicated that the culture of S. nudus in tidal flat will not result in significant deterioration of sediment. The OM and fine sand of the five layers in the P group increased significantly than those in the S and C groups, which indicated that the Meretrix lyrata had strong filter feeding and deposition ability, and the bioturbation of S. nudus can accelerate the migration of OM and fine sand. The ORP value of the two layers (0~3, 3~10 cm) in the P group decreased significantly than that in the S group (p<0.05), and the results indicated that the high density of Meretrix lyrata will result in significant deterioration of their habitat. S. nudus can migrate down and bury surface organic matter, and M. lyrata can promote the deposition of particulate matter; the complementary functions of the two animals help improve the nutrient utilization and carbon sequestration function of the tidal flat.
Key words: Sipunculus nudus Linnaeus; Meretrix lyrata; culturing; sediment
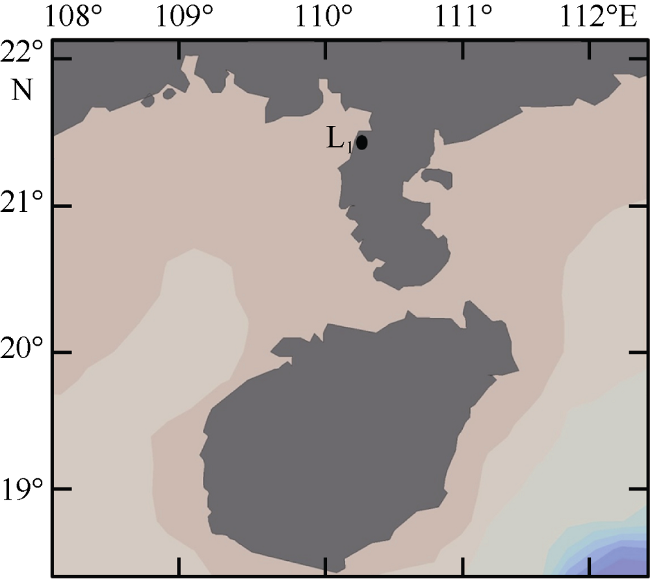
Fig. 1 Location of the studying area in Beibu Gulf (L1)图1 北部湾方格星虫增养殖区及调查地点(L1) |
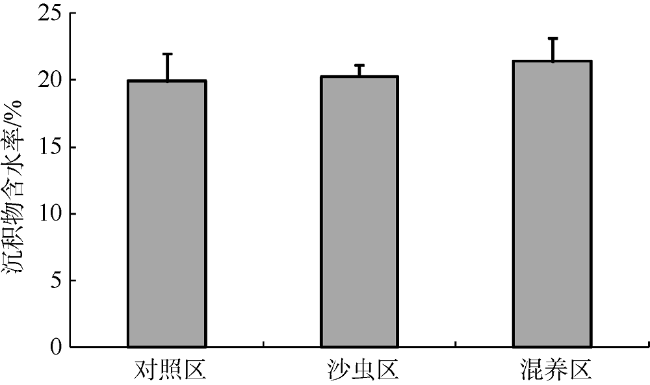
Fig. 2 The water content of tidal flat sediment in each of the three groups图2 不同增养殖区滩涂底质含水率 |
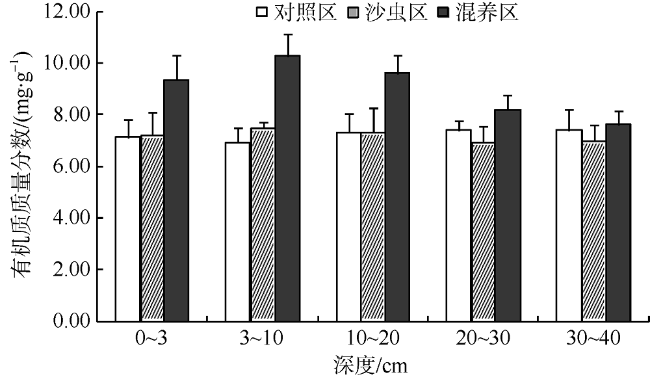
Fig. 3 Organic matter content in different layers in spring图3 春季不同层次沉积物中的有机质质量分数 |
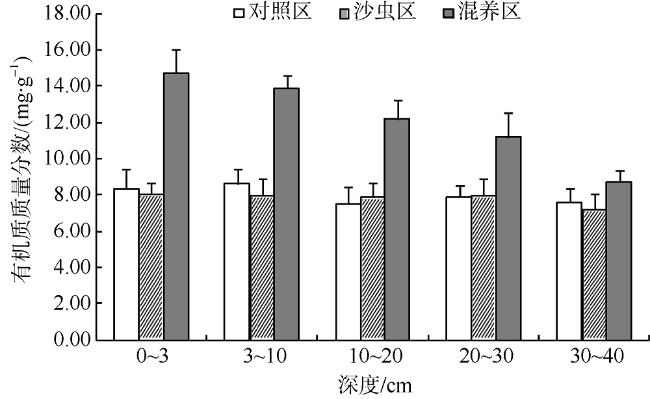
Fig. 4 Organic matter content in different layers in autumn图4 秋季不同层次沉积物中的有机质质量分数 |
Fig. 5 The oxidation reduction potential (ORP) of different layers in spring图5 春季不同层次沉积物中的氧化还原电位值变化 |
Fig. 6 The oxidation reduction potential of different layers in autumn图6 秋季不同层次沉积物中的氧化还原电位值变化 |
Tab. 1 The grain composition of tidal flat sediment in spring表1 春季不同增养殖区滩涂底质粒径组成 |
| 粒径组成 | 处理组 | 0~3cm | 3~10cm | 10~20cm | 20~30cm | 30~40cm |
|---|---|---|---|---|---|---|
| d>0.85mm 沙粒占比/% | 对照区 | 38.62±3.36 | 23.21±3.02 | 23.68±3.18 | 29.27±3.62 | 31.39±2.89 |
| 沙虫区 | 23.26±1.69 | 25.03±2.33 | 26.93±3.26 | 33.23±2.59 | 34.29±3.65 | |
| 混养区 | 22.56±2.56 | 23.68±2.69 | 27.38±3.02 | 32.16±3.23 | 32.31±2.98 | |
| 0.85mm>d>0.43mm 沙粒占比/% | 对照区 | 20.32±3.02 | 26.58±2.21 | 29.59±2.37 | 30.79±4.03 | 32.92±2.61 |
| 沙虫区 | 38.57±4.23 | 36.38±3.69 | 35.59±2.13 | 35.56±5.32 | 37.75±3.26 | |
| 混养区 | 36.52±4.31 | 35.61±4.10 | 34.13±3.41 | 36.57±4.67 | 37.66±4.09 | |
| 0.43mm>d>0.18mm 沙粒占比/% | 对照区 | 31.53±3.18 | 38.76±4.35 | 37.25±2.72 | 31.13±3.02 | 29.09±2.69 |
| 沙虫区 | 29.39±3.04 | 32.69±3.52 | 30.52±2.61 | 24.31±3.68 | 21.94±2.23 | |
| 混养区 | 32.41±3.08 | 33.26±2.06 | 29.31±2.83 | 26.41±3.76 | 24.23±2.51 | |
| 0.18mm>d>0.1mm 沙粒占比/% | 对照区 | 7.63±1.02 | 9.36±1.69 | 8.12±1.15 | 6.31±0.96 | 4.91±1.35 |
| 沙虫区 | 6.84±0.69 | 4.57±0.93 | 4.68±0.52 | 4.23±0.38 | 4.18±0.25 | |
| 混养区 | 5.92±0.81 | 4.23±0.62 | 4.92±0.12 | 4.13±0.64 | 4.68±0.58 | |
| d<0.1mm 沙粒占比/% | 对照区 | 0.65±0.13 | 1.32±0.20 | 0.83±0.09 | 1.69±0.16 | 1.36±0.08 |
| 沙虫区 | 0.63±0.05 | 0.52±0.08 | 0.65±0.11 | 0.68±0.14 | 0.85±0.20 | |
| 混养区 | 0.71±0.18 | 0.56±0.13 | 0.59±0.09 | 0.71±0.07 | 0.80±0.09 |
Tab. 2 The grain composition of tidal flat sediment in autumn表2 秋季不同增养殖区滩涂底质粒径组成 |
| 粒径组成 | 处理组 | 0~3cm | 3~10cm | 10~20cm | 20~30cm | 30~40cm |
|---|---|---|---|---|---|---|
| d>0.85mm 沙粒占比/% | 对照区 | 44.88±6.37 | 17.37±3.21 | 22.90±2.36 | 32.59±4.09 | 33.21±2.67 |
| 沙虫区 | 29.40±3.01 | 25.35±2.15 | 23.83±3.41 | 25.44±3.96 | 24.18±2.91 | |
| 混养区 | 28.29±4.64 | 17.66±1.82 | 16.26±2.13 | 28.35±3.64 | 27.62±3.04 | |
| 0.85>d>0.43mm 沙粒占比/% | 对照区 | 16.91±2.06 | 25.77±3.19 | 30.86±2.82 | 30.85±3.17 | 31.03±4.53 |
| 沙虫区 | 30.75±4.13 | 33.28±2.27 | 34.45±4.61 | 33.11±2.98 | 33.12±4.91 | |
| 混养区 | 26.06±3.83 | 25.54±2.61 | 26.72±2.37 | 30.37±3.64 | 29.06±3.17 | |
| 0.43>d>0.18mm 沙粒占比/% | 对照区 | 29.49±4.13 | 44.11±5.26 | 37.80±4.01 | 28.37±3.28 | 27.21±3.94 |
| 沙虫区 | 34.00±5.13 | 35.81±2.69 | 35.05±4.37 | 35.62±4.16 | 36.19±5.38 | |
| 混养区 | 34.69±2.16 | 41.87±5.64 | 41.94±4.28 | 33.08±3.01 | 34.59±4.53 | |
| 0.18>d>0.1mm 沙粒占比/% | 对照区 | 7.85±1.02 | 11.24±1.85 | 7.63±0.52 | 5.75±0.63 | 4.23±0.41 |
| 沙虫区 | 5.36±0.68 | 5.12±0.81 | 6.07±1.03 | 5.15±0.59 | 5.26±0.67 | |
| 混养区 | 8.17±1.21 | 11.61±1.06 | 12.22±1.39 | 6.90±0.81 | 7.15±0.79 | |
| d<0.1mm 沙粒占比/% | 对照区 | 0.87±0.19 | 1.50±0.08 | 0.81±0.05 | 2.45±0.27 | 1.67±0.31 |
| 沙虫区 | 0.49±0.05 | 0.45±0.06 | 0.59±0.12 | 0.68±0.09 | 0.65±0.11 | |
| 混养区 | 2.79±0.31 | 3.33±0.26 | 2.86±0.32 | 1.29±0.17 | 1.35±0.25 |
Tab. 3 Diatom density in surface sediment in autumn表3 秋季滩涂沉积物表层的硅藻密度(单位: ×104个•cm-2) |
| 组别 | 对照区 | 沙虫区 | 混养区 |
|---|---|---|---|
| 针杆藻Synedra sp. | 85 | 102 | 17 |
| 粗针杆藻Synedra Robusta Ralfs | 50 | 17 | 34 |
| 舟形藻Navicula sp. | 170 | 85 | 34 |
| 双壁藻Diploneis sp. | - | - | 17 |
| 卵形藻Cocconeis sp. | - | - | 17 |
| 舟形藻 斑点亚属Navicula Punctatae sp. | - | - | 17 |
| 总数 | 305 | 204 | 136 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.6-2007 海洋调查规范第6部分: 海洋生物调查[S]. 北京: 中国标准出版社.
GENERAL ADMINISTRATION OF QUALITY SUPERVISION, INSPECTION AND QUARANTINE OF THE PEOPLE’S REPUBLIC OF CHINA, STANDARDIZATION ADMINISTRATION OF THE PEOPLE’S REPUBLIC OF CHINA, 2008. GB/T 12763.6-2007, Specifications for oceanographic survey Part 6: Marine biological survey[S]. Beijing: Standards Press of China (in Chinese).
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}