
两种造礁石珊瑚对海水酸化和溶解有机碳加富的响应*
作者简介:郭亚娟(1990—), 女, 河南省周口市人, 硕士, 从事海洋环境生态学研究。E-mail: guoyajuan15@mails.ucas.ac.cn
收稿日期: 2017-02-17
要求修回日期: 2017-06-16
网络出版日期: 2018-02-02
基金资助
国家自然科学基金(31370499、31370500)
国家科技支撑(2014BAC01B03)
Responses of two species of reef-building corals to acidification and dissolved organic carbon enrichment*
Received date: 2017-02-17
Request revised date: 2017-06-16
Online published: 2018-02-02
Supported by
National Natural Science Foundation of China Projects (31370499, 31370500)
National Key Technology Support Program (2014BAC01B03)
Copyright
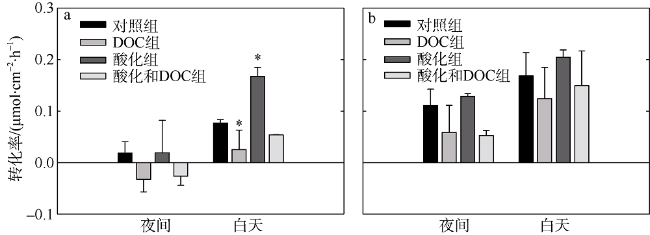
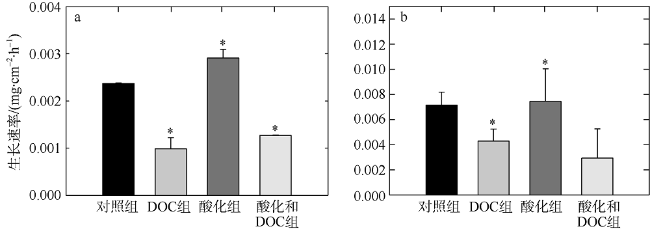
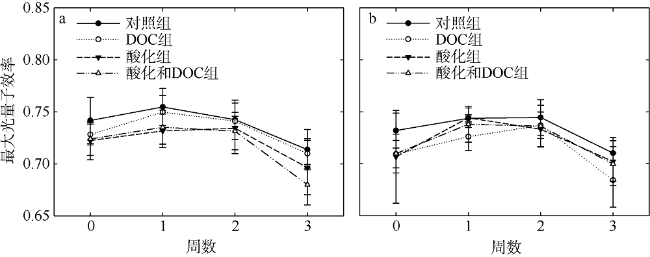
文章选择鹿回头近岸海域常见的板叶角蜂巢珊瑚(Favites complanata)和十字牡丹珊瑚(Pavona decussata)为研究对象, 采用室内连续培养的方法, 探究两种不同造礁石珊瑚对酸化和溶解有机碳(DOC)加富的响应。结果表明: 酸化(pH 7.6)并不会影响两种珊瑚的钙化速率和生长速率; 但DOC加富(524.03±78.42μmol•L-1)使两种珊瑚的钙化速率分别降低67%和47%、生长速率降低59%和40%。当二者共同作用时, 两种珊瑚的钙化速率降低30%和11%、生长速率降低46%和59%, 大多没有DOC单独作用时强烈, 表现出一定的拮抗作用。两种珊瑚共生虫黄藻叶绿素荧光指数(Fv/Fm)均升高后降低, 板叶角蜂巢珊瑚Fv/Fm最先降低。实验表明, 这两种珊瑚虽然对海洋酸化的敏感度不高, 但是对有机物加富有不同的响应, 板叶角蜂巢珊瑚更为敏感, 可能导致这两种珊瑚在未来环境变化中有不同命运。
郭亚娟 , 周伟华 , 袁翔城 , 廖健祖 , 江雷 , 黄晖 . 两种造礁石珊瑚对海水酸化和溶解有机碳加富的响应*[J]. 热带海洋学报, 2018 , 37(1) : 57 -63 . DOI: 10.11978/2017018
Two common reef-building coral species in Luhuitou offshore areas, Favites complanata and Pavona decussata, were selected in our experiments. Continuous culture experiments (21 days) were conducted to evaluate the effects of acidification and dissolved organic carbon (DOC) enrichment on coral physiology. The results showed that: 1) acidification (pH 7.6) did not significantly affect the calcification rates and growth rates of these two coral species. However, in the treatment of DOC enrichment (524.03±78.42μmol•L-1), the coral calcification rates decreased by 67% and 47%, respectively; the growth rates decreased by 59% and 40%, respectively. In the combined effects of acidification and organic matter enrichment, the calcification rates decreased by 67% and 47%, respectively; the growth rates decreased by 59% and 40%, respectively, but the reduction was less than DOC enrichment alone. To some degree, these results reflected some antagonistic action. The maximum quantum efficiency (Fv/Fm) of these two corals all increased in the early, meanwhile; Fv/Fm of Favites complanata decreased first. 2) Favites complanata and Pavona decussata responded differently to acidification and organic matter enrichment, indicating the species would have different fates in a changing environment in future.
Fig. 1 Two species of reef-building corals. (a) Favites complanata; and (b) Pavona decussate图1 两种造礁石珊瑚 |
Tab. 1 pH & DOC treatments表1 pH和DOC处理 |
| 组别 | pH | 溶解有机碳浓度/(μmol•L-1) |
|---|---|---|
| 对照组 | 8.1 | 153.83±6.33 |
| DOC组 | 8.1 | 524.03±78.42 |
| 酸化组 | 7.6 | 153.83±6.33 |
| 酸化和DOC组 | 7.6 | 524.03±78.42 |
Fig. 2 Effects of acidification and DOC enrichment on the calcification rates of Favites complanata and Pavona decussate. (a) Favites complanata; (b) Pavona decussata图2 酸化和DOC加富对板叶角蜂巢珊瑚(a)和十字牡丹珊瑚(b)钙化速率的影响 |
Fig. 3 Effects of acidification and DOC enrichment on the growth rates of Favites complanata and Pavona decussata. (a) Favites complanata; (b) Pavona decussate图3 酸化和DOC加富对板叶角蜂巢珊瑚(a)和十字牡丹珊瑚(b)生长速率的影响 |
Fig. 4 Effects of acidification and DOC enrichment on the photosynthesis capacity (Fv/Fm) of Favites complanata and Pavona decussata. (a) Favites complanata; (b) Pavona decussate图4 酸化和DOC加富对板叶角蜂巢珊瑚(a)和十字牡丹珊瑚(b)光合能力(Fv/Fm)的影响 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}