
缢蛏多巴胺受体基因及其在损伤修复中的作用
作者简介:杜蕴超(1993—), 女, 山东省威海市人, 硕士, 研究方向: 贝类种质资源与遗传育种。E-mail:dorayunc@qq.com
收稿日期: 2017-06-09
要求修回日期: 2018-01-02
网络出版日期: 2018-05-03
基金资助
国家自然科学基金项目(31472278)
国家高技术研究发展计划(2012AA10A400-3)
Dopamine receptor genes of Sinonovacula constricta and its functions in damage healing of tissue
Received date: 2017-06-09
Request revised date: 2018-01-02
Online published: 2018-05-03
Supported by
National Basic Research Program of China (31472278)
National High-tech R & D Program (“863” Program) of China (2012AA10A400-3)
Copyright
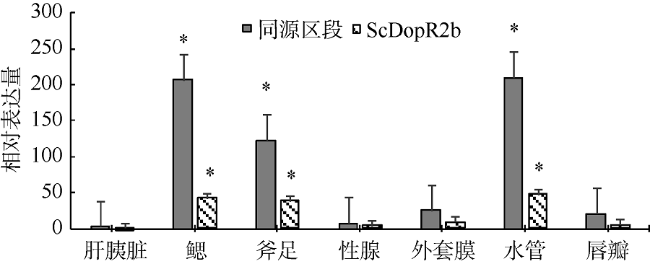
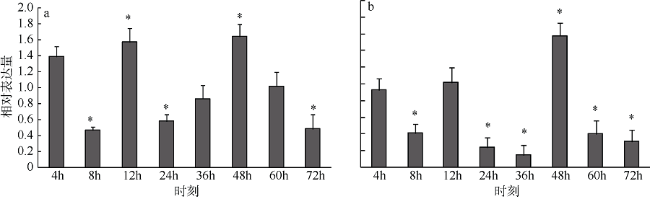
多巴胺受体(dopamine receptor)作为生物体中重要的神经递质受体, 在生物体的生长、发育、代谢等多个生理过程中都发挥着重要的功能。本研究从缢蛏Sinonovacula constricta转录组文库中筛选获得多巴胺受体基因的部分片段, 结合RACE(rapid-amplification of cDNA ends)技术和降落 PCR(polymerase chain reaction)技术, 克隆得到缢蛏两个多巴胺受体的选择性多聚腺苷酸化(alternative polyadenylation, APA)的变异体, 命名为ScDopR2a和 ScDopR2b, 长度分别为1824bp和2758bp。两个变异体均包含相同的5′非翻译区(untranslated regions, UTR)(24bp)和开放阅读框(open reading frame, ORF)(1440bp), 共编码479个氨基酸残基。但是两个变异体的3′UTR 长度不同, 分别为360bp和1294bp, 其中ScDopR2b在polyA尾前插入936bp。在同源的ORF区设计引物, 采用实时定量PCR分析多巴胺受体基因在不同组织中的表达特征, 结果表明多巴胺受体基因在水管、鳃、斧足的表达量显著高于其他组织。在该基因序列3′UTR插入片段区域设计特异引物检测ScDopR2b的组织表达情况, 结果表明在水管、鳃、斧足的表达量仍高于其他组织, 表达趋势与ORF区大致相同。进一步设计缢蛏组织损伤实验, 对进水管前端进行损伤处理, 在处理后的第4h、8h、12h、24h、48h、60h和72h取样, 荧光定量检测结果显示, 同源区表达量在12h和48h呈上调, 在8h、24h和72h下调, 且ScDopR2b表达趋势与同源区表达模式大致相同。研究结果表明, 缢蛏多巴胺受体参与了损伤修复过程, 其表达特征可能与多巴胺作为补偿性神经递质的作用有关。
杜蕴超 , 谢淑媚 , 何圣耀 , 牛东红 , 李家乐 . 缢蛏多巴胺受体基因及其在损伤修复中的作用[J]. 热带海洋学报, 2018 , 37(3) : 45 -54 . DOI: 10.11978/2017068
Dopamine receptor is an important neurotransmitter receptor, playing a key role in organism growth, development, metabolism, and other physiological processes. In this study, we obtained a fragment of dopamine receptor gene from transcriptome library of razor clam (Sinonovacula constricta), and cloned the full-length cDNA of dopamine receptor gene using RACE (rapid-amplification of cDNA ends) and Touchdown PCR (polymerase chain reaction). The full-length cDNA sequence contains two isoforms of alternative polyadenylation (APA), named as ScDopR2a and ScDopR2b, with a length of 1824 bp and 2758 bp, respectively. Both isoforms contain a 24 bp 5′untranslated region (UTR), and a 1440 bp open reading frame encoding 479 amino acid residues. However, the 3′UTR length is 360 bp and 1294 bp for ScDopR2a and ScDopR2b, respectively, with an insertion of 936 bp in front of the polyA tail for ScDopR2b. Primers were designed in open reading frame (ORF)to detect the expression level of dopamine receptor gene (ScDopR2) by quantitative real-time PCR. The results revealed the expression in waterpipe, gill and foot were higher than that in the other tissues. Then, primers were designed in 3′UTR to detect the expression level of ScDopR2b, showing similar expression pattern with ORF region. In the tissue damage experiment, the samples were selected at 4, 8, 12, 24, 48, 60, and 72 h after injuring waterpipe. The fluorescence quantitative results showed that a significant increase in 12 and 48 h, and a significant decrease in 8, 24 and 72 h. The trend of ScDopR2b gene’s expression level was similar to that of ORF. The results suggested that dopamine receptors are involved in the damage-healing process, and the expression pattern may be due to a role of dopamine as compensatory neurotransmitters.
Tab. 1 Information of primers used for PCR analysis表1 引物序列信息 |
| 引物 | 序列(5′-3′) | 用途 |
|---|---|---|
| F | ATCCACAGTGCTACAAGAGTTGC | 中间片段扩增 |
| R | CTGCTGACGACGTCTGTTCT | 中间片段扩增 |
| 3R1 | AGTGAAAGTGACGAGGACGACCATACCAAC | 3′RACE |
| 3R2 | TTCCAACCACGAGGTGCAAACAGAAGCTAC | 3′RACE |
| 3R3 | ATCAACATCACAACAATCAGCGACCAATG | 3′RACE |
| 3R4 | TTCATCACCAAACGTCTCCGACAATTAGC | 3′RACE |
| 3R5 | TACAGATTCTACCAGAACAGACGTCGTCAGC | 3′RACE |
| 下游内部引物 | CGCGGATCCTCCACTAGTGATTTCACTATAGG | 3′RACE |
| 下游外部引物 | TACCGTCGTTCCACTAGTGATTT | 3′RACE |
| 18SF | TCGGTTCTATTGCGTTGGTTTT | 荧光定量内参引物 |
| 18SR | CAGTTGGCATCGTTTATGGTCA | 荧光定量内参引物 |
| ScdoprF | CTTCCAATCGGCTGTTCG | ORF区段荧光定量引物 |
| ScdoprR | GGTATGGTCGTCCTCGTCAC | ORF区段荧光定量引物 |
| ScDopR2bF | AACTGTCCGTAACGCCAAAG | ScDopR2b特异区荧光定量引物 |
| ScDopR2bR | TCAAACATCCCACTTCGGG | ScDopR2b特异区荧光定量引物 |
| ORF-F | CATCACAACAATCAGCGAC | 半定量验证引物 |
| ORF-R | ACAACAATCCCAAGCGTC | 半定量验证引物 |
| UTR-F | TTCAGCGTGGTTGATGTG | 半定量验证引物 |
| UTR-R | CTTTGGCGTTACGGACAGT | 半定量验证引物 |
注: F表示上游引物; R表示下游引物。 |
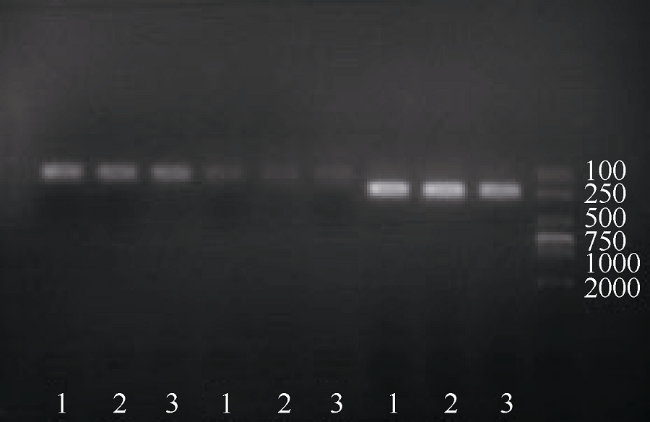
Fig. 1 The cloning picture of two isoforms of ScDopR2 gene from S. constricta图 1 缢蛏ScDopR2基因两个多聚腺苷酸化变异体克隆电泳图 |
Fig. 2 The semi-quantitative PCR picture of two isoforms of ScDopR2 gene from S. constricta图 2 缢蛏ScDopR2基因两个多聚腺苷酸化变异体半定量电泳图 |
Fig. 3 Full-length of cDNA and deduced amino acid sequence of ScDopR2 gene from S. constricta图 3 缢蛏ScDopR2编码基因cDNA全长及推导的氨基酸序列 |
Tab. 2 Prediction information of alternative polyadenylation signal of ScDopR2 gene from S. constricta表 2 缢蛏ScDopR2基因选择性多聚腺苷酸加尾信号的预测信息 |
| 位置/bp | PAS序列 | 分值 |
|---|---|---|
| 1720 | TATAAA | 0.469 |
| 1727 | TATAAA | 0.577 |
| 1794 | TATAAA | 0.558 |
| 2735 | AATAAA | 0.097 |
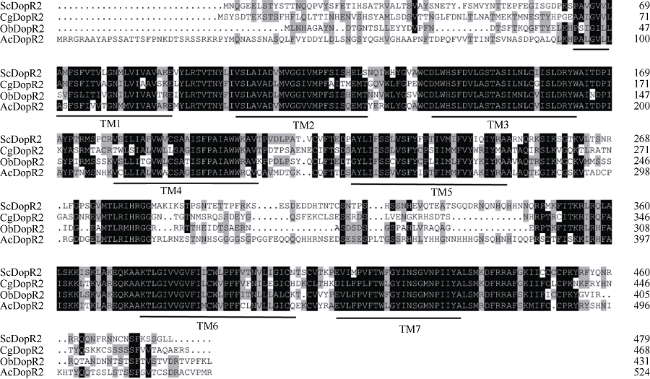
Fig. 4 Multiple alignment of the amino acid sequence of ScDopR2 with its homologues in other organisms图 4 缢蛏和其他物种的DopR2基因氨基酸序列的多重比对 |
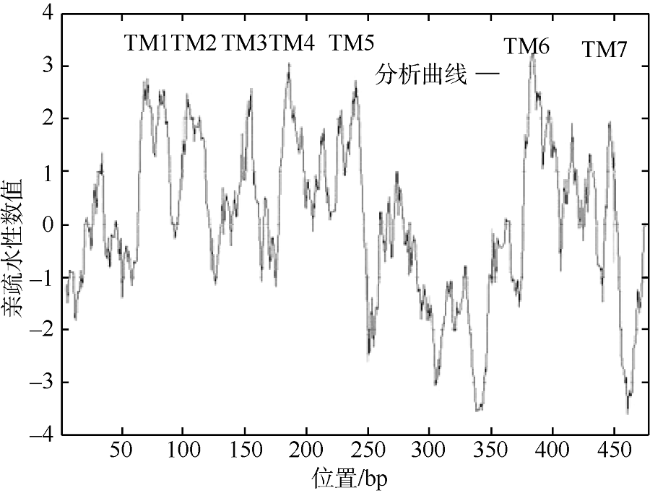
Fig. 5 Hydrophobicity analysis of ScDopR2 protein from S. constricta (by Protscale)图 5 缢蛏ScDopR2编码蛋白质的疏水性分析(由Protscale程序预测) |
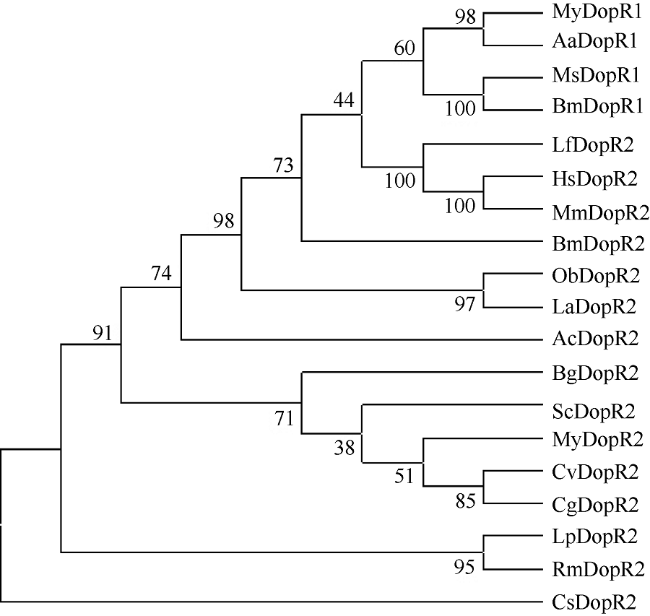
Fig. 6 Maximum-Parsimony phylogenetic tree of dopamine receptor amino acid sequences图 6 基于MP法构建的多巴胺受体基因系统进化树 |
Tab. 3 Information of the sequences used in the phylogenetic analysis表 3 用于系统进化分析的序列信息 |
| 基因名 | 物种名 | 登录号 |
|---|---|---|
| AaDopR1 | Aedes aegypti | AFB73766.1 |
| BmDopR1 | Bombyx mori | NP_001108459.1 |
| MyDopR1 | Mizuhopecten yessoensis | XP_021361185.1 |
| MsDopR1 | Manduca sexta | AEU17117.1 |
| LfDopR2 | Lampetra fluviatilis | ADO23655.3 |
| HsDopR2 | Homo sapiens | NP_000786.1 |
| MyDopR2 | Mizuhopecten yessoensis | XP_021340507.1 |
| BgDopR2 | Biomphalaria glabrata | XP_013063747.1 |
| CvDopR2 | Crassostrea virginica | XP_022345762.1 |
| CgDopR2 | Crassostrea gigas | XP_019919825.1 |
| AcDopR2 | Aplysia californica | XP_005099999.1 |
| RmDopR2 | Rhipicephalus microplus | AFC88980.1 |
| CsDopR2 | Chilo suppressalis | AKR18179.1 |
| MmDopR2 | Mus musculus | NP_034207.2 |
| BmDopR2 | Bombyx mori | NP_001108338.1 |
| ObDopR2 | Octopus bimaculoides | XP_014768037.1 |
| LaDopR2 | Lingula anatina | XP_013419306.1 |
| LpDopR2 | Limulus polyphemus | XP_013783833.1 |
Fig. 7 mRNA expression level of dopamine receptor gene homology region and ScDopR2b from S. constricta图 7 缢蛏多巴胺受体基因同源区段和 ScDopR2b mRNA的组织表达 |
Fig. 8 The expression level of dopamine receptor homology region and ScDopR2b from S. constricta in waterpipe damage图 8 缢蛏多巴胺受体基因同源区段和ScDopR2b在组织损伤中的表达 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}