
深海来源微生物乙酰酯酶的酶学性质鉴定及拆分制备D-乳酸甲酯
作者简介:黄锦龙(1986—), 男, 博士研究生, 研究方向为生物催化与酶工程。E-mail: jl_huang@163.com
收稿日期: 2017-09-29
要求修回日期: 2017-10-25
网络出版日期: 2018-07-16
基金资助
中国科学院战略性先导科技专项项目(XDA11030404);广东省海洋渔业科技攻关与研发方向项目(A201701C12);广州市科技计划项目(201510010012)
Characterization of one deep-sea derived microbial acetyl esterase and its utilization in the preparation of D-methyl lactate through kinetic resolution
Received date: 2017-09-29
Request revised date: 2017-10-25
Online published: 2018-07-16
Supported by
Strategic Priority Research Program of the Chinese Academy of Sciences (XDA11030404);Scientific and Technological Project of Ocean and Fishery from Guangdong Province (A201701C12);Guangzhou Science and Technology Plan Projects (201510010012)
Copyright
D-乳酸及其酯是重要的手性药物中间体和手性化工产品。从南海深海芽孢杆菌Bacillus sp. SCSIO15029克隆到一个乙酰酯酶基因bae02030, 表达并鉴定该酶Bae02030的酶学性质。该酯酶的最适pH和最适温度分别为8.5和35℃, 其对多种有机溶剂和表面活性剂具有较好的耐受性。乙酰酯酶Bae02030能够通过水解拆分消旋乳酸甲酯来制备光学纯的D-乳酸甲酯。通过对拆分反应进行优化, 添加体积分数为60%的正庚烷能够改善乙酰酯酶Bae02030的光学选择性, 所制备的D-乳酸甲酯的对映体过量值(e.e.s)超过99%, 转化率(c)为56%。深海微生物来源的乙酰酯酶Bae02030作为生物催化剂在工业上制备手性药物中间体具有较好的应用潜力。
黄锦龙 , 张继福 , 胡洁莹 , 关见留 , 张云 , 孙爱君 , 胡云峰 . 深海来源微生物乙酰酯酶的酶学性质鉴定及拆分制备D-乳酸甲酯[J]. 热带海洋学报, 2018 , 37(4) : 38 -44 . DOI: 10.11978/2017106
D-lactic acid and its esters are important chiral drug intermediates and chemicals. An acetyl esterase, Bae02030, from Bacillus sp. SCSIO15029 isolated from the deep sea of the South China Sea was cloned, expressed and functionally characterized. The optimum pH and temperature of acetyl esterase Bae02030 were 8.5 and 35°C, respectively. Bae02030 exhibited excellent resistance to most organic solvents and surfactants tested. Bae02030 could generate optically pure D-methyl lactate through resolution of racemic methyl lactate. After process optimization of the enzymatic resolution reactions, the addition of 60% heptane improved the enantio-selectivity of Bae02030. The enantiomeric excess of prepared D-methyl lactate was over 99% and conversion rate reached 56%. Acetyl esterase Bae02030 identified from deep-sea microorganism, as a biocatalyst, possesses great potential in the production of chiral drug intermediates in industry.
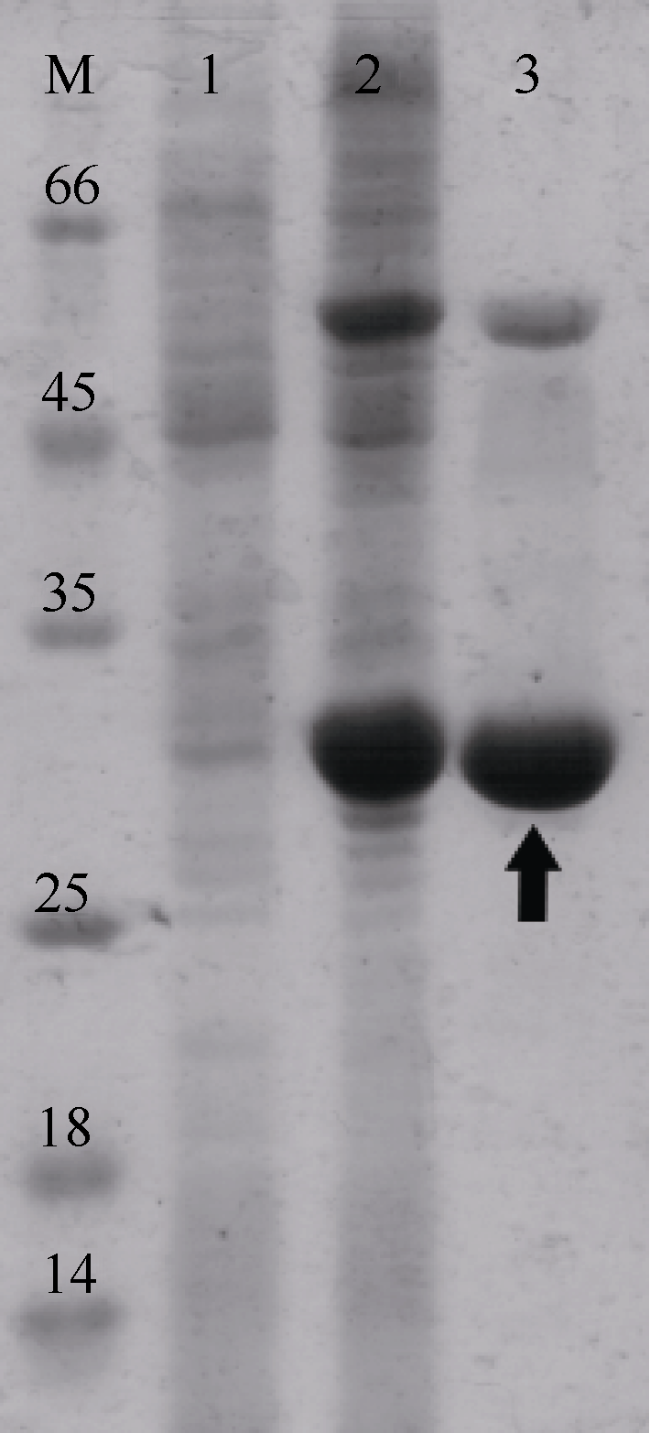
Fig. 1 SDS-PAGE of Bae02030. M. markers; 1. total proteins of recombinant stain before IPTG induction; 2. total proteins of recombinant stain after IPTG induction; 3. purified Bae02030图1 Bae02030蛋白SDS-PAGE电泳 |
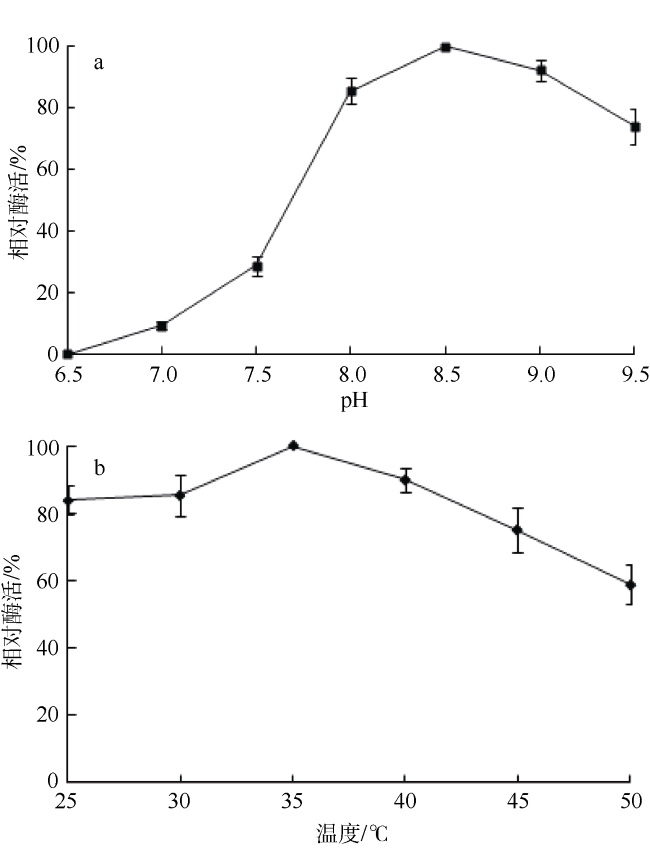
Fig. 2 Effects of pH (a) and temperature (b) on Bae02030 activity图2 pH (a)和温度(b)对Bae02030活性的影响 |
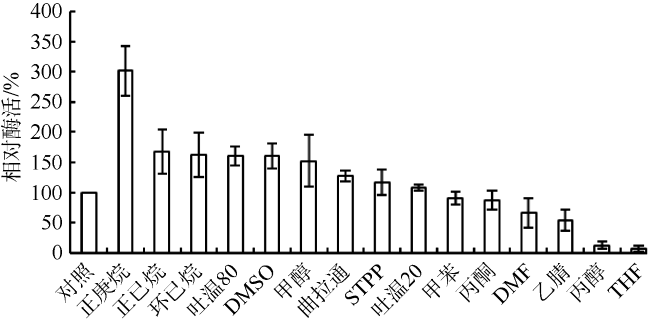
Fig. 3 Effects of organic solvents and surfactants on the relative activity of Bae02030图3 有机溶剂和表面活性剂对Bae02030酶活的影响 |
Tab. 1 Effects of pH and temperature on the resolution of methyl lactate by Bae02030表1 pH和温度对Bae02030拆分乳酸甲酯的影响 |
| pH | 对映体 过量值/% | 转化率/% | 温度/°C | 对映体 过量值/% | 转化率/% |
|---|---|---|---|---|---|
| 7.5 | 35.85 ± 0.31 | 53.73 ± 0.25 | 30 | 71.17 ± 0.44 | 55.29 ± 0.40 |
| 8.0 | 48.13 ± 0.38 | 55.0 ± 0.30 | 35 | 76.33 ± 0.28 | 56.42 ± 0.18 |
| 8.5 | 66.05 ± 0.62 | 56.23 ± 0.29 | 37 | 81.17 ± 0.35 | 57.86 ± 0.51 |
| 9.0 | 76.45 ± 0.53 | 56.96 ± 0.52 | 40 | 83.29 ± 0.53 | 58.47 ± 0.32 |
| 9.5 | 65.21 ± 0.33 | 58.21 ± 0.41 | 45 | 79.87 ± 0.71 | 60.08 ± 0.33 |
| 10.0 | 63.79 ± 0.19 | 59.31 ± 0.38 | 50 | 64.79 ± 0.36 | 59.57 ± 0.59 |
Tab. 2 Effects of organic solvents and surfactants on the resolution of methyl lactate by Bae02030表2 有机溶剂和表面活性剂对Bae02030拆分乳酸甲酯的影响 |
| lgP | 对映体过量值/% | 转化率/% | |
|---|---|---|---|
| 对照 | 0 | 89.23 ± 0.42 | 66.60 ± 0.55 |
| 正己烷 | 3.94 | 89.15 ± 0.27 | 66.76 ± 0.32 |
| 环己烷 | 3.39 | 90.27 ± 0.46 | 66.32 ± 0.42 |
| 正庚烷 | 4.47 | 90.53 ± 0.52 | 61.34 ± 0.20 |
| 甲醇 | -0.72 | 52.89 ± 0.63 | 54.97 ± 0.46 |
| 甲苯 | 2.68 | 84.23 ± 0.35 | 64.60 ± 0.34 |
| 丙酮 | -0.16 | 50.19 ± 0.30 | 45.88 ± 0.09 |
| DMSO | -1.35 | 88.89 ± 0.73 | 61.94 ± 0.24 |
| 曲拉通 | 0 | 88.23 ± 0.33 | 67.30 ± 0.22 |
| 吐温-20 | 0 | 90.53 ± 0.25 | 69.80 ± 0.88 |
| 吐温-80 | 0 | 91.33 ± 0.41 | 66.42 ± 0.27 |
| STPP | 0 | 90.15 ± 0.95 | 67.57 ± 1.04 |
注: lgP为有机化合物脂水分配系数, 数值参考自www. chemspider.com |
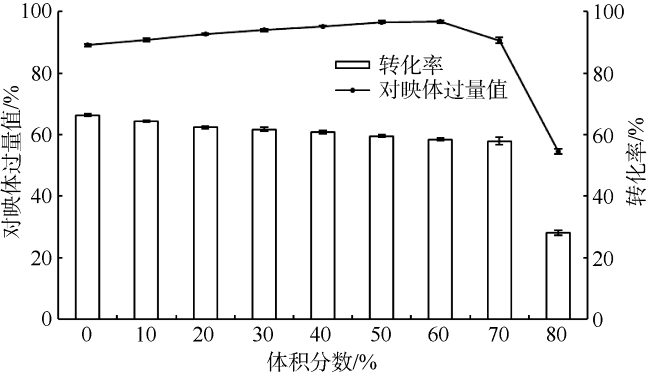
Fig. 4 Effect of concentration of heptane on the resolution by Bae02030图4 不同正庚烷体积分数对Bae02030拆分的影响 |
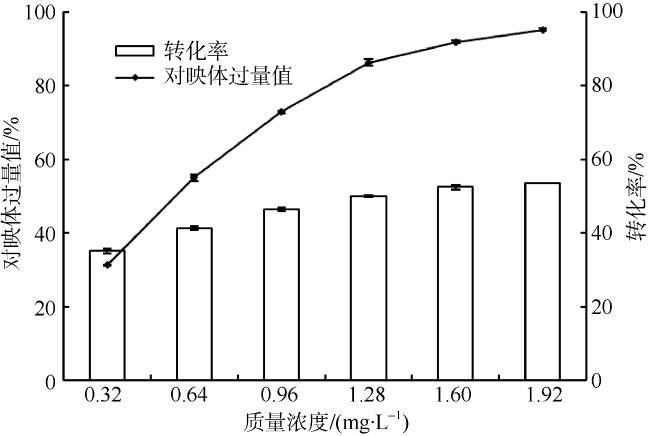
Fig. 5 Effects of enzyme loading on the resolution of methyl lactate图5 酶的质量浓度对拆分反应的影响 |
Tab. 3 Effects of different substrate concentrations on the resolution of methyl lactate by Bae02030表3 不同底物浓度对Bae02030拆分乳酸甲酯的影响 |
| 底物浓度 /(mmol·L-1) | 时间/h | 对映体 过量值/% | 转化率/% | D-乳酸甲酯产率 /(mmol·L-1·h-1) |
|---|---|---|---|---|
| 12.5 | 2 | > 99.5 | 56.03 ± 0.43 | 2.75 ± 0.03 |
| 25.0 | 4 | > 99.5 | 61.43 ± 0.26 | 2.41 ± 0.03 |
| 37.5 | 6 | > 99.5 | 63.35 ± 0.35 | 2.29 ± 0.04 |
| 50.0 | 6 | 87.07 ± 0.31 | 54.58 ± 0.17 | 1.78 ± 0.01 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}