
南海东北部夏季叶绿素a浓度垂向变化特征及其对水动力过程的响应*
作者简介: 徐文龙(1991—), 男, 山东省枣庄市人, 硕士研究生, 主要从事海洋光学研究。E-mail: wenlongxu@scsio.ac.cn
收稿日期: 2017-11-10
要求修回日期: 2018-03-07
网络出版日期: 2018-10-13
基金资助
国家自然科学基金(41776045、41376042、41576030、41776044)
热带海洋环境国家重点实验室自主研究项目(LTOZZ1602)
Vertical variability of chlorophyll a concentration and its responses to hydrodynamic processes in the northeastern South China Sea in summer
Received date: 2017-11-10
Request revised date: 2018-03-07
Online published: 2018-10-13
Supported by
National Natural Science Foundation of China (41776045, 41376042, 41576030, 41776044)
Open Project Program of the State Key Laboratory of Tropical Oceanography (South China Sea Institute of Oceanology, Chinese Academy of Sciences) (LTOZZ1602)
Copyright
叶绿素a浓度是估算海洋初级生产力的一个重要参数, 在海洋中垂向分布极不均匀, 其分布特征及影响机制是海洋生态学研究的重要基础问题。利用海洋光学观测的高垂向分辨率剖面数据, 系统地研究叶绿素a浓度垂向剖面的时空分布特征及其与海洋动力环境要素的关系, 可为揭示南海典型动力过程的生态环境效应提供重要基础。文章基于2015年夏季黑潮调查航次实测生物光学剖面, 利用676nm处吸收基线高度(aLH(676))与叶绿素a浓度(Chla)之间的关系, 建立了具有较高反演精度的叶绿素a浓度反演算法(Chla=49.96×(aLH(676))0.9339, 决定系数R2=0.87和均方根误差RMSE=0.16mg·m-3); 进一步结合观测期间物理过程, 揭示了叶绿素垂向分布对不同水动力过程的响应特征。研究结果表明, 近岸区域表层叶绿素a浓度变化范围为0.42~1.57mg·m-3, 随着水深增加, 叶绿素a浓度逐渐降低, 在沿岸上升流影响区域, 叶绿素a浓度明显增高, 垂向上相对趋于均一分布; 次表层叶绿素极大值(Subsurface Chlorophyll Maximum, SCM)现象在外海显著存在, 受中尺度过程影响明显, SCM深度在34m到100m之间变化, 在吕宋岛以西海域, 黑潮入侵加速了上层水体的混合, SCM所在水层被显著抬升至34m左右; 在冷涡影响区域, 次表层叶绿素极大值层被抬升, 涡旋中心比涡旋边缘抬升更为显著, 同时SCM的厚度增大。
徐文龙 , 王桂芬 , 周雯 , 许占堂 , 曹文熙 . 南海东北部夏季叶绿素a浓度垂向变化特征及其对水动力过程的响应*[J]. 热带海洋学报, 2018 , 37(5) : 62 -73 . DOI: 10.11978/2017121
Chlorophyll a (Chla) concentration is an important indicator of phytoplankton biomass for estimating primary production, which exhibits inhomogeneous vertical distribution. In this study, optical measurements with high vertical resolution were used for studying the spatio-temporal variability of Chla profiles and its influencing mechanism, providing us much insights for understanding marine environmental dynamics in the South China Sea. In this study, an optical inversion method was proposed to retrieve Chla profiles from in situ measurements with high-spectral absorption/attenuation spectrophotometer in the northeastern South China Sea. Based on the in situ data of the Kuroshio Cruise in summer 2015, an empirical relationship between the absorption line height at 676 nm and Chla was developed for inverting Chla with a high accuracy (Chla=49.96×(aLH(676) (676))0.9339, the coefficient of determination R2=0.87, the root mean square error RMSE=0.16 mg·m-3). Moreover, the response characteristics of Chla profiles to different hydrodynamic processes were studied. Results showed that large spatial differences existed in the vertical distribution of Chla. In the nearshore area, surface Chla varied from 0.42 to 1.57 mg·m-3, which generally followed a decreasing trend with depth. Uniform vertical distribution of Chla in upper ocean was observed in the coastal upwelling region. The Subsurface Chlorophyll Maximum (SCM) layer is nearly ubiquitous in stratified waters of the open ocean, with its vertical structure being influenced by mesoscale processes. The depth of SCM ranged between 34 and 100 m, which showed consistent variability with the isopycnal depth of 1023 kg·m-3. The SCM layer was uplifted to about 34 m due to the mixing effect in the upper ocean caused by Kuroshio intrusion in the western area of Luzon Island. The uplifting and widening of the SCM layer were also observed in the area affected by a cold eddy, with obviously different features in the eddy center from that on the eddy edge.
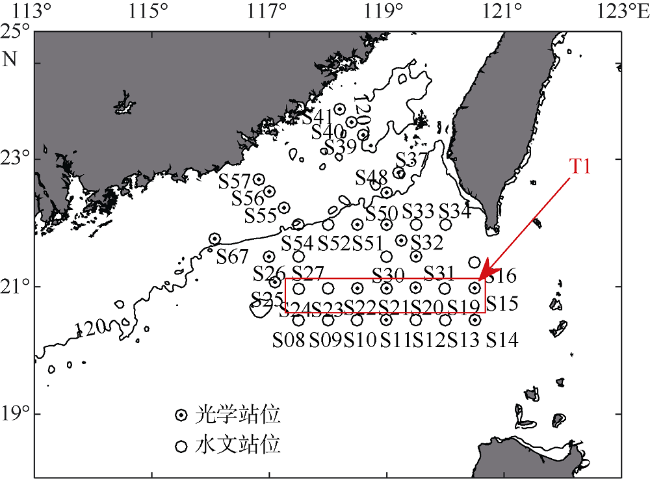
Fig. 1 Sampling sites in the northern South China Sea during August 2015. The section profile (T1) used in |
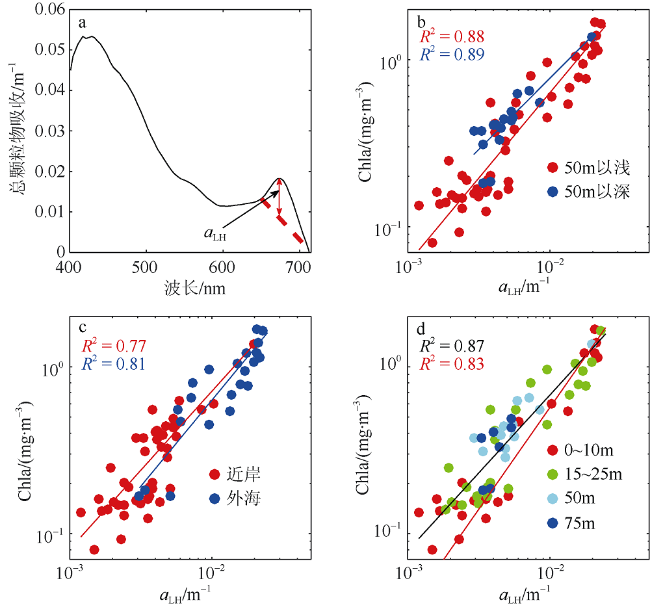
Fig. 2 Estimation of Chlorophyll a concentration from a bio-optical algorithm. The description of absorption line height (a) and scatter plots of the absorption line height in 676 nm and chlorophyll a concentration, for a variety of standards of division (b, c, d). (d) Black line is the regression obtained in this study, while the red line is based on Boss et al (2013). The coefficient of determination between the logarithms of aLH(676) and Chla is 0.87, and the root mean square error (RMSE) is 0.16 mg·m-3. N=69图2 叶绿素a浓度生物光学反演算法 |
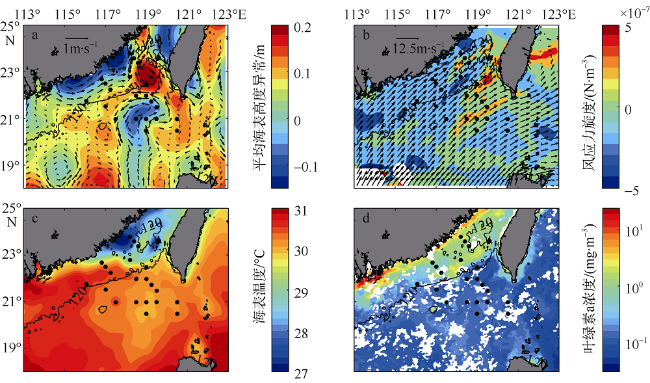
Fig. 3 Spatial distributions of mean sea level anomaly and geostrophic velocities (a), sea surface wind and wind stress curl (b), and sea surface temperature (c), together with sea surface chlorophyll during the survey (d). All data are downloaded from the websites (MSLA: ftp.aviso.altimetry.fr; SSW: ftp://eclipse.ncdc.noaa.gov/pub/seawinds/; SST: http://www.ghrsst-pp.org; Chla: http://hermes. acri.fr). Vectors in (a) and (b) are surface geostrophic currents and wind speed, respectively. Geostrophic current is from the AVSIO data. MSLA, SeaWind, SST, and Chla are 13-day-mean data during the period 5-17 August 2015. Wind stress curl is the data of 7 August 2015图3 航次期间南海北部遥感数据分布 |
Fig. 4 Chlorophyll a profiles estimated from absorption line height. Red/black solid lines represent stations with water depth lower/higher than 120 m图4 基于吸收线性高度反演得到的叶绿素垂向分布剖面 |
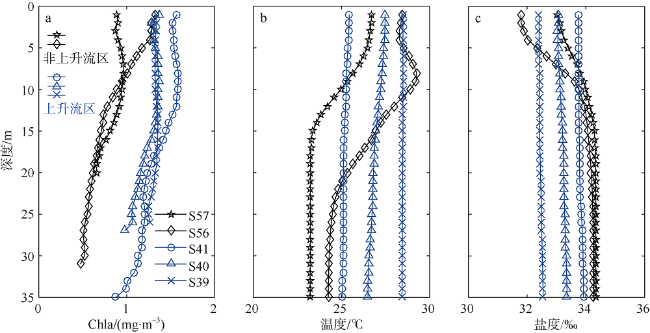
Fig. 5 Vertical distributions of chlorophyll a concentration (a), temperature (b) and salinity (c). Blue and red are marked inside and outside the upwelling influenced area, respectively图5 近岸叶绿素a浓度(a)、温度(b)和盐度(c)及分布剖面 |
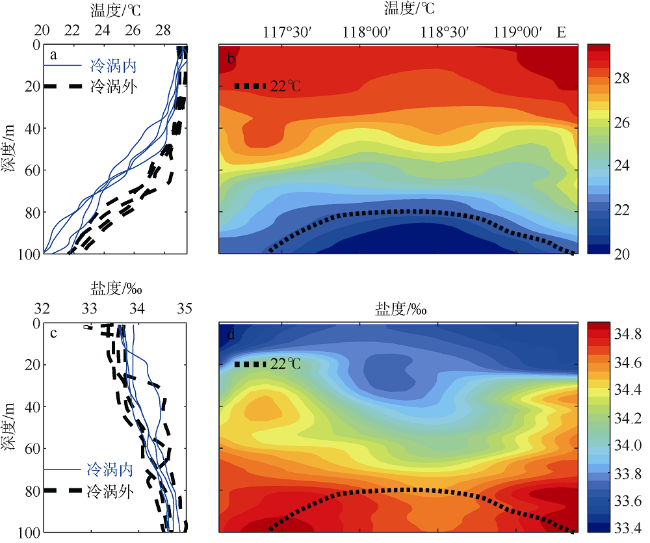
Fig. 6 Temperature (a) and salinity (b) profiles within the cold eddy. Sectional distribution of temperature (c) and salinity (d) along transect T1. The black dashed line corresponds to the thermocline (defined as the 22°C contour)图6 冷涡断面(T1)的温度及盐度分布 |
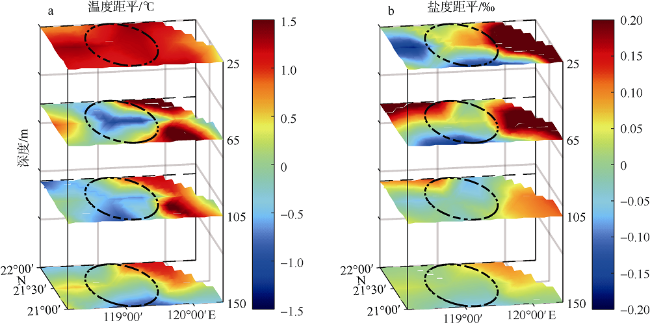
Fig. 7 Three-dimensional structures of temperature anomaly (°C)(a) and salinity anomaly (‰)(b). The black dashed line indicates the area affected by the cold eddy图7 冷涡影响范围温度距平(°C) (a)和盐度距平(‰)切面图(b) |
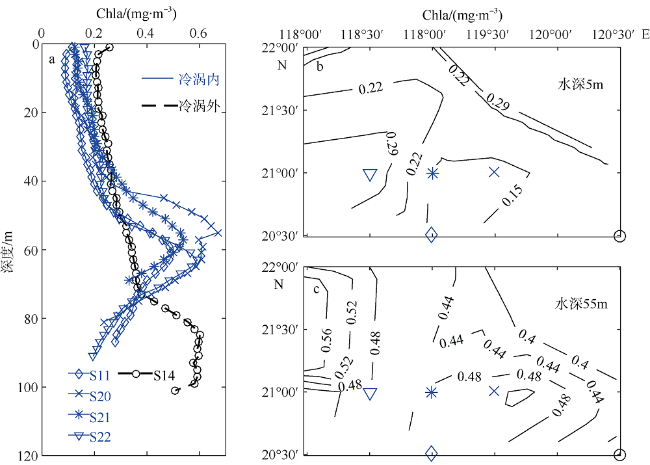
Fig. 8 Vertical distribution of chlorophyll a concentration (mg·m-3) in a cold eddy in (a). Horizontal distribution of chlorophyll a concentration (mg·m-3) in a cold eddy (b, c); the contour intervals in (b) and (c) are 0.07 mg·m-3 and 0.04 mg·m-3, respectively. The symbols represent the station information used in (a)图8 冷涡叶绿素a浓度(mg·m-3)分布剖面及切面 |
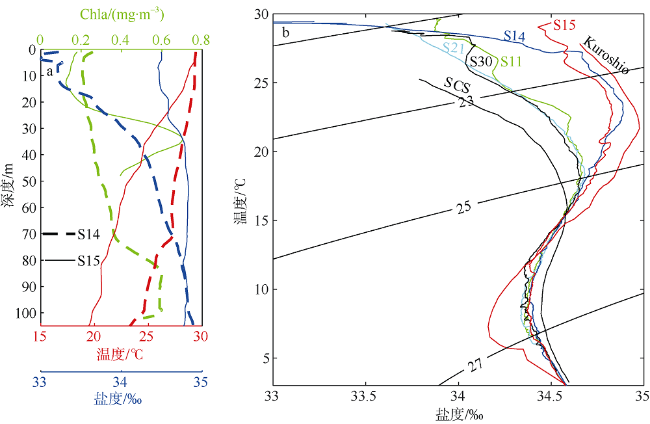
Fig. 9 Observed chlorophyll a concentration (green), temperature (red) and salinity (blue) profiles at the two sites in the western area of Luzon Island (a) and temperature versus salinity plots for five stations in the western area of Luzon Island (b). Typical South China Sea (SCS) and Kuroshio T/S curves are based on measurements made by Chen and Huang (1996). Solid lines are isopycnal contour lines (units: kg·m-3 ). S11, S20 and S21 are located in the western Luzon Strait; S14 and S15 are close to the region of Kuroshio intrusion图9 黑潮影响区叶绿素a浓度及温盐分布 |
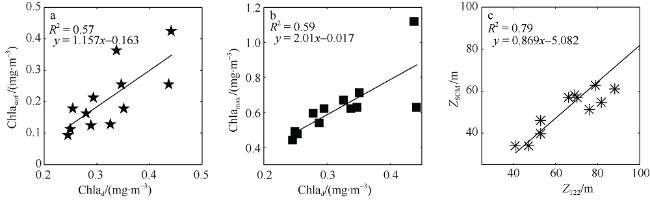
Fig. 10 Statistical analysis of related parameters. Chlad: The average of water column integrated chlorophyll a concentration;, Chlamax: subsurface chlorophyll maximum; ZSCM: subsurface chlorophyll maximum layer; ZT22: depth of 22°C; N=12图10 相关参数统计分析图 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
SHAN,
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}