
季风环流影响下的南海海洋细菌多样性特征初探
作者简介: 李洁(1983—), 女, 湖北省英山县人, 研究员, 博士, 海洋微生物学。E-mail: lijietaren@scsio.ac.cn
收稿日期: 2017-11-29
网络出版日期: 2018-12-24
基金资助
国家自然科学基金(41230962)
Preliminary investigation on the bacteria diversity coupled with the monsoon forced circulation in the South China Sea
Received date: 2017-11-29
Online published: 2018-12-24
Supported by
National Natural Science Foundation of China (41230962)
Copyright
季风驱动下的南海海洋动力过程有着季节性的多尺度变化特征, 显著影响南海海洋生态系统的演化进程。海洋细菌作为海洋生态系统中物质循环和能量流动的重要组成部分, 对环境变化和海洋多尺度动力过程有着积极的响应。将微生物生态过程与海洋动力过程相结合, 研究微生物的群落结构、动态变化及其与海洋环境过程的耦合, 是目前国际海洋科学多学科交叉研究的热点之一。文章以国家自然科学基金重点项目“季风环流影响下的南海海洋细菌多样性特征及其生物海洋学意义”的主要研究成果为基础, 从南海北部上升流、海洋锋面、中尺度涡旋、次中尺度过程及其对海洋细菌多样性和生态系统可能的影响等几个方面, 探讨当前的研究进展和初步研究成果。

李洁 , 经志友 , 张偲 . 季风环流影响下的南海海洋细菌多样性特征初探[J]. 热带海洋学报, 2018 , 37(6) : 1 -15 . DOI: 10.11978/2017127
Ocean dynamic processes in the South China Sea driven by the monsoon are characterized by seasonal and multi-scale variations, which have significant impacts on the evolution of marine ecosystems. Marine bacteria, as an important component of material cycle and energy flow in marine ecosystems, have active responses to the environmental changes associated with multi-scale dynamic processes. The interdisciplinary research on microbial community structures, their dynamic variation and associated physical processes is one of the hot issues in marine sciences, which combines marine microbiology, microbial ecology and physical oceanography. Based on the primary results of the Key Research Program, "Study on bacteria diversity coupled with the monsoon circulation in the South China Sea and significance in microbial oceanography" supported by the National Natural Science Foundation of China, we discuss in this paper current research progress, including coastal upwelling, seasonal thermal fronts, mesoscale eddy, sub-mesoscale processes, and their potential impacts on the marine bacterial diversity and local ecosystem in the South China Sea.
Fig. 1 Typical circulations in the South China Sea in winter (top) and summer (bottom), using AVISO altimeter data |
Fig. 2 Satellite observation (a); numerical simulation (b); in-situ observations (c, d), and analysis of water masses (e) for upwelling in the northern South China Sea. |
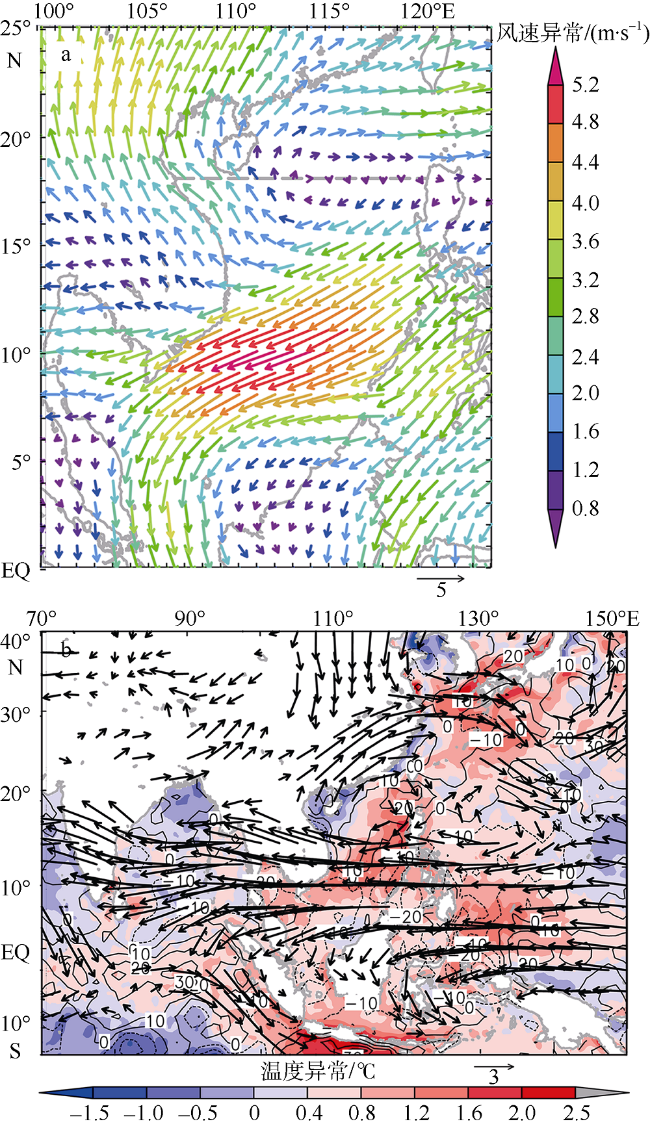
Fig. 3 South China Sea monsoon anomaly (a); atmospheric circulation anomalies in the western Pacific, South China Sea and East Indian Ocean (b) in the summer of 1998. |
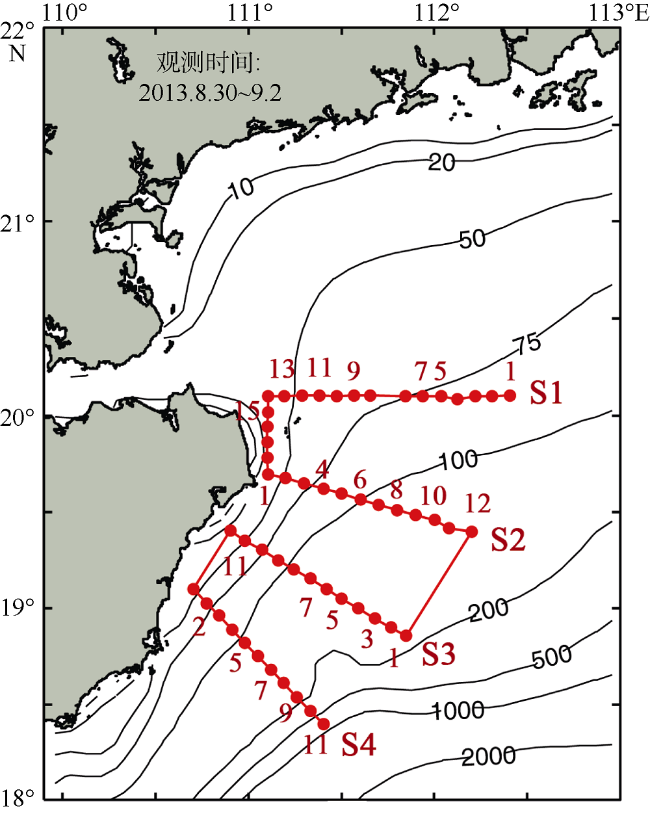
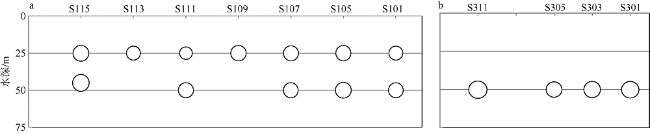
Fig. 4 Map of water depths overlapped with CTD stations (dots). |
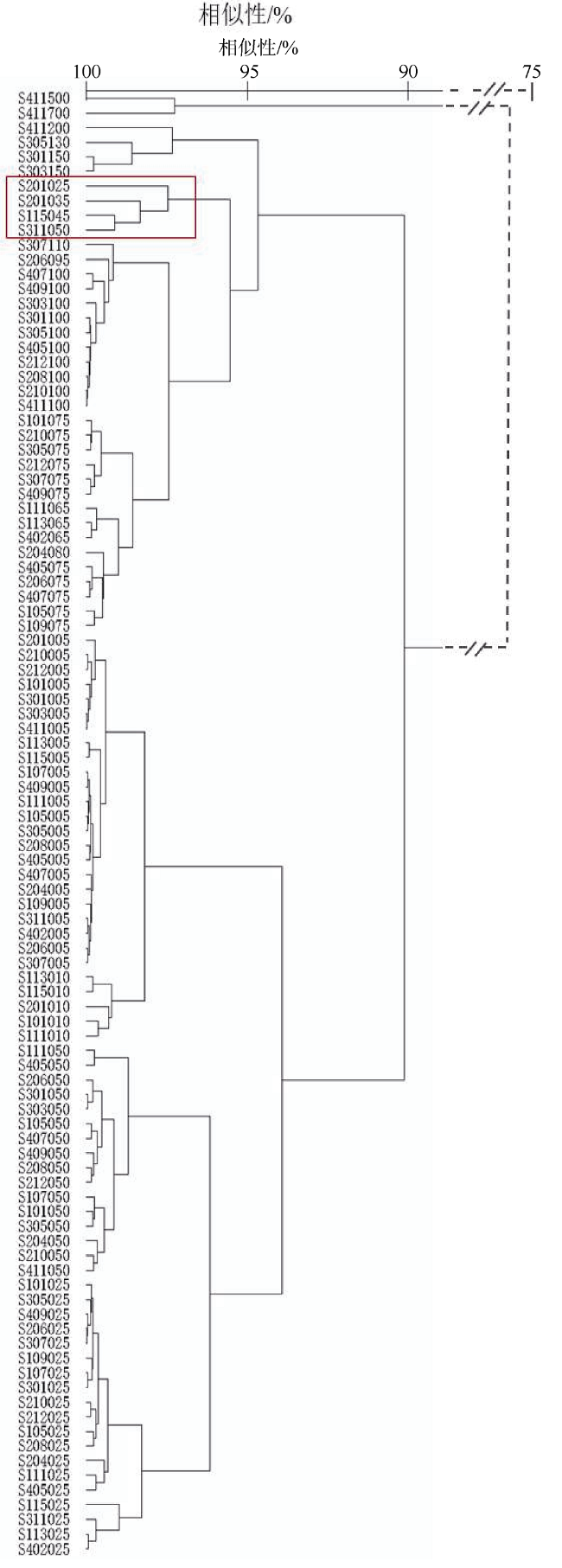
Fig. 5 Cluster analysis of the environmental elements at the sampling stations. |
Fig. 6 Bacterial diversity at 25 and 50 m along transect S1 (a) and at 50 m along transect S3 (b). |
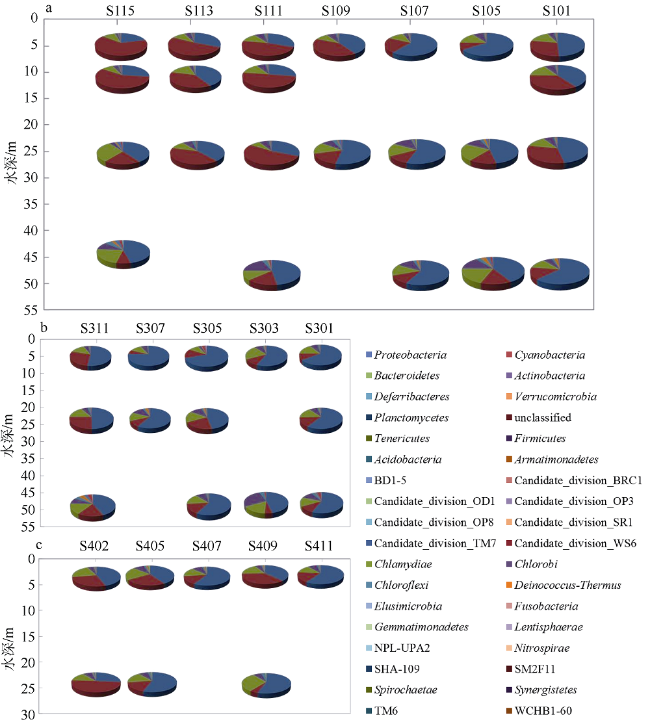
Fig. 7 Bacterial community compositions at different depths of the sampling stations |
Fig. 8 Annual-mean SST gradients and wind speeds from Satellite observations (a); in-situ observations (b) and vertical structures of temperature and salinity in the representative sections (c-h) for the fronts in the northern South China Sea. |
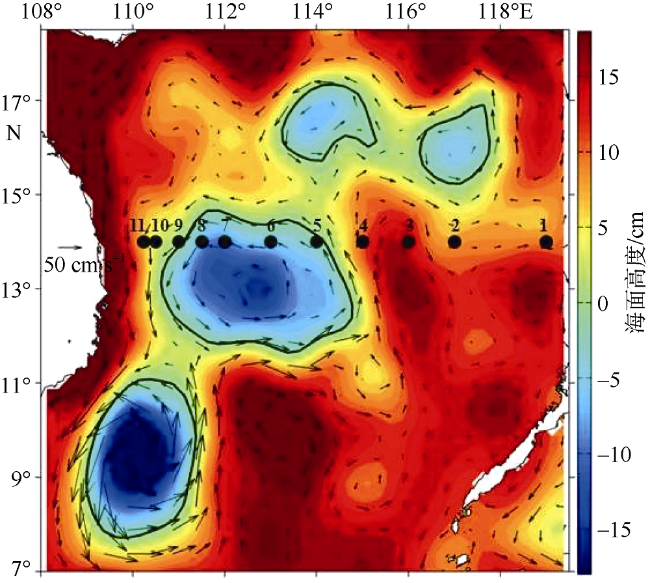
Fig. 9 Distributions of sea level anomalies and geostrophic velocity anomalies during the cruise, overlapped with sampling stations |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}