
夏季南海北部浮游植物对上升流与羽状流的响应
作者简介:许泽婷(1993—), 女, 广东省广州市人, 硕士研究生, 主要从事近岸海域生态环境模拟研究。E-mail: xuzeting@mail2.sysu.edu.cn
收稿日期: 2018-01-01
网络出版日期: 2018-12-24
基金资助
中央高校基本科研业务费专项资金(17lgzd20);热带海洋环境国家重点实验室(中国科学院南海海洋研究所)开放课题(LTO1605);国家自然科学基金(41306105);广东省自然科学基金(2014A030313169);
Summer phytoplankton responses to upwelling and river plume in northern South China Sea
Received date: 2018-01-01
Online published: 2018-12-24
Supported by
Fundamental Research Funds for the Central Universities (17lgzd20);State Key Laboratory of Tropical Oceanography, South China Sea Institute of Oceanology, Chinese Academy of Sciences (LTO1605);National Natural Science Foundation of China (41306105);National Natural Science Foundation of Guangdong Province (2014A030313169)
Copyright
文章建立了基于真实场驱动的三维物理—生态耦合模型, 利用模型定量分析了夏季南海北部上升流和羽状流过程对浮游植物生物量空间分布的影响程度及作用机制。首先, 利用2006—2008年卫星遥感数据及2006与2008年夏季观测数据对模型进行了验证, 结果表明, 模型能较好地再现夏季南海北部上升流和羽状流过程, 较好地反映出浮游植物的空间分布特征。模拟分析结果显示, 夏季南海北部浮游植物主要分布在50m等深线以内。琼州海峡东部海域和汕头海域浮游植物垂向分布较为均匀, 上升流的贡献均达到90%以上, 表层水平平流输送是浮游植物主要的汇, 生物过程是浮游植物的源。珠江口和汕尾海域浮游植物存在表层和次表层两个高值区, 羽状流贡献35%~40%, 主要促进表层浮游植物生长, 而上升流贡献60%~65%, 主要促进中底层浮游植物的生长。粤西海域上升流对浮游植物的贡献占92%, 主要促进中底层浮游植物生长, 而表层浮游植物浓度极低。整体上, 夏季南海北部上升流和羽状流主要是通过输送营养盐的方式影响浮游植物的生长。上升流对营养盐的输送作用是向岸方向的爬升输送和平行于等深线的沿岸流输送共同作用的结果。跃层的存在改变了营养盐的垂向输送过程, 是导致上升流和羽状流过程对不同水层浮游植物贡献差异的关键因素之一。整体而言, 夏季南海北部浮游植物空间分布差异是以上升流、羽状流主导, 环流—营养盐—生物过程共同作用的结果。
关键词: 物理—生态耦合模型;; 浮游植物; 上升流; 羽状流; 南海北部
许泽婷 , 李适宇 , 胡嘉镗 , 王思颖 , 汪斌 , 郭铭先 , 耿兵绪 . 夏季南海北部浮游植物对上升流与羽状流的响应[J]. 热带海洋学报, 2018 , 37(6) : 92 -103 . DOI: 10.11978/2018001
A coupled physical-biological 3D model was established, which is forced by realistic forcing, to quantify the effects and underlying mechanisms of upwelling and river plume on the spatial distribution of phytoplankton biomass during summer in the northern South China sea (NSCS). The model was validated by using cruise data of 2006 and 2008 and remote-sensing observations from 2006 to 2008. The results suggest that the model satisfactorily captures the processes of coastal upwelling and river plume dynamics that determine phytoplankton distribution. Model results indicate that summer phytoplankton in the NSCS were mainly distributed within 50m isobath. In Qiongdong and Shantou, phytoplankton distributed evenly. Upwelling processes accounted for up to 90% of the biomass, and horizontal advection in the upper layer was the main sink, while biological processes were the main sources for the phytoplankton biomass. In the Pearl River and Shanwei, phytoplankton shows surface and subsurface maxima: the river plume contributed 35%~40% (mainly to the upper layer) of the phytoplankton biomass, and upwelling contributed 60%~65% (mainly to the middle and bottom layers). In Yuexi, the phytoplankton biomass was extremely low in the upper layer, and mainly distributed in the middle and bottom layers; in total, upwelling contributed 92% of phytoplankton biomass. In the NSCS, both upwelling and river plume processes stimulated phytoplankton through nutrient supply. In the upwelling process, nutrient supply reflected the combination of along- and across-shore nutrient transports. The thermocline, which alters vertical nutrient transfer, is the key factor affecting the variable contributions of upwelling and river plume processes for phytoplankton in different layers. Overall, the spatial variation of summer phytoplankton is mainly driven by upwelling and river plume processes, and by the circulation-nutrient-biological coupling effect.
Fig. 1 (a) Model Calculation area (black box) and water depth. |
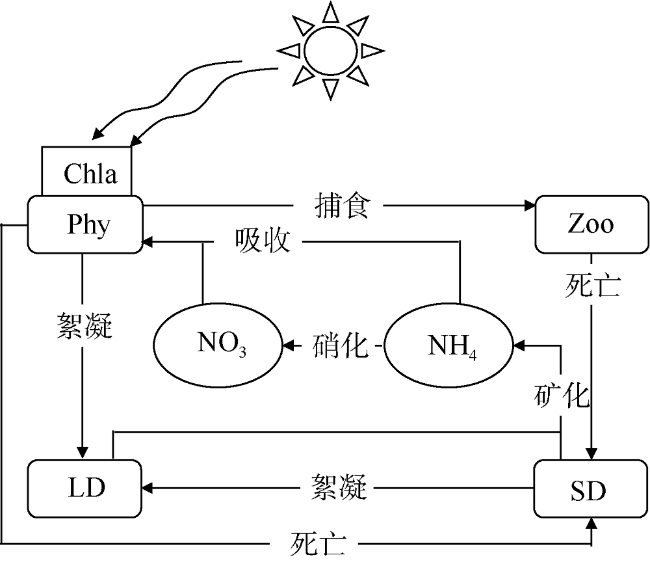
Fig. 2 Schematic of the biological model图2 NPZD生态模型生物地球化学反应过程示意 |
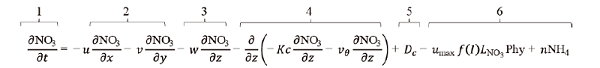 (1)
(1)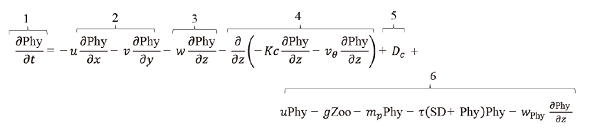 (2)
(2)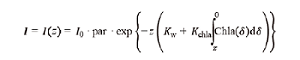 (3)
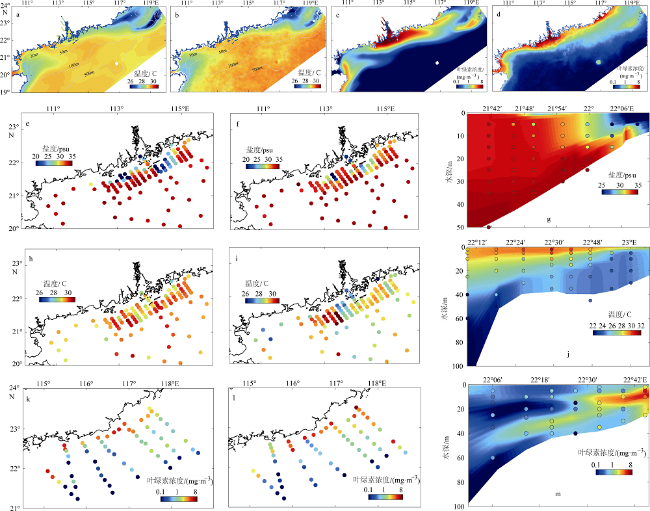
(3)Fig. 3 Comparison of simulated and observed of temperature (℃), salinity (psu) and chlorophyll (mg·m-3)图3 表层温度、盐度和叶绿素浓度模拟值与观测值的比较 |
Tab. 1 Validation metrics of the physical-biological coupling model表1 水动力-生态耦合模型验证统计指标 |
| 验证指标 | 2006—2008年遥感数据 | 2006年大面站 | 2008年大面站 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bias | RMSE | R | RE/% | Bias | RMSE | R | RE/% | Bias | RMSE | R | RE/% | |
| 海表温度 | -0.02 | 0.28 | 0.90 | 0.06 | 1.03 | 0.52 | 0.39 | 1.77 | 0.54 | 1.18 | 0.84 | 1.99 |
| 海表盐度 | — | — | — | — | 3.56 | -1.51 | 0.85 | 4.73 | 0.59 | 2.04 | 0.60 | 1.86 |
| 海表叶绿素 | -0.13 | 1.08 | 0.75 | 17.92 | — | — | — | — | -0.25 | 1.78 | 0.46 | 14.88 |
注: “—”表示缺测; Bias表示平均偏差(计算平均值-观测平均值, 温度单位为℃, 盐度单位为psu, 叶绿素浓度单位为mg·m-3); RMSE表示均方根误差, 单位与Bias一致; R表示模拟值与观测值地相关性系数; RE表示相对平均误差。 |
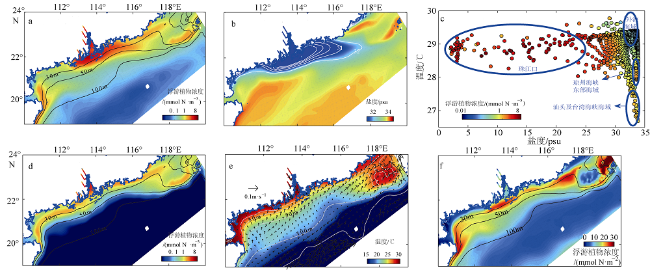
Fig. 4 Summertime surface, bottom and vertically-integrated physical parameters and phytoplankton distribution图4 夏季表层、底层及垂向积分浮游植物及水动力分布 |
Fig. 5 Phytoplankton, temperature and velocity, and NO3 distributions for sections L1~L5图5 L1—L5断面浮游植物、温度、流场和NO3分布 |
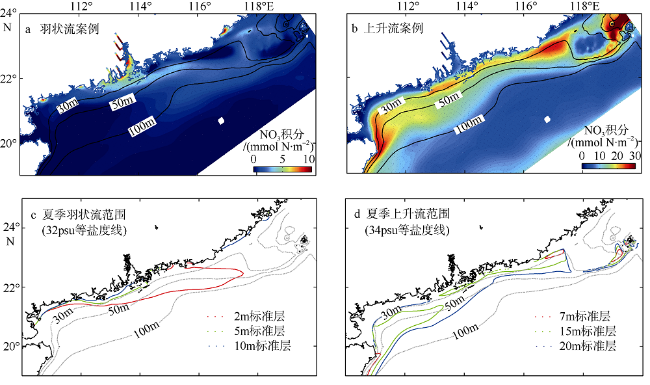
Fig. 6 Vertically-integrated phytoplankton concentration (a, b); upwelling and river plume range (c, d)图6 夏季浮游植物垂向积分浓度的贡献图(a、b)及上升流与羽状流的范围(c、d) |
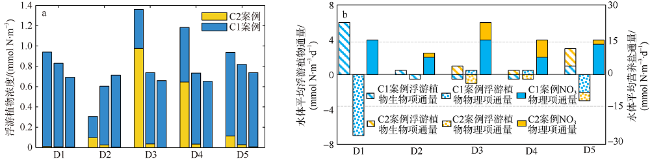
Fig. 7 (a) Upper, middle and bottom layer phytoplankton concentrations in sub-regions. (b) Diagnostic results in sub-regions图7 不同区域C1、C2案例的上、中、底层浮游植物浓度(a)及各区域水体平均诊断项结果(b) |
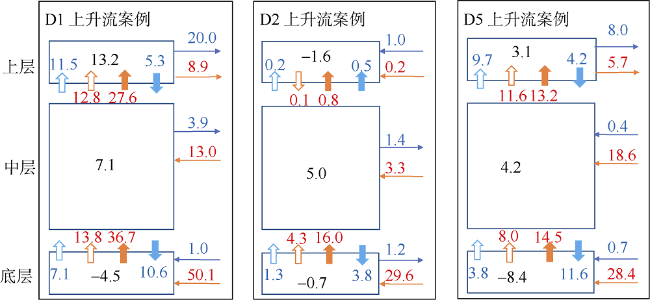
Fig. 8 Phytoplankton and NO3 diagnostic results for sub-regions D1, D2 and D5 in case C1图8 D1、D2和D5区域在C1案例中浮游植物及NO3收支 |
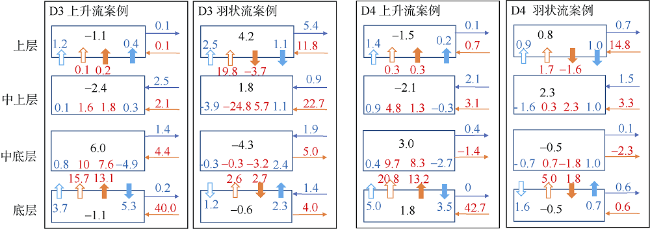
Fig. 9 Same as |
Fig. 10 Vertically-integrated NO3 flux for across (a) and along (b) seashore directions for case C1图10 C1案例水体积分跨岸(a)及沿岸(b)方向NO3通量图 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.7-2007 海洋调查规范第7部分: 海洋调查资料交换[S]. 北京: 中国标准出版社.
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}