
Shewanella oneidensis内源性大质粒上ParE-ParD家族毒素-抗毒素系统(SO_A0088-A0087)的鉴定
作者简介:赵玄玉(1993—), 女, 广东省珠海市人, 海洋生物学硕士, 从事毒素-抗毒素系统的功能研究。E-mail: zhaoxuanyu88@163.com
收稿日期: 2018-02-28
网络出版日期: 2018-12-24
基金资助
国家自然科学基金(31500025、31500093)
Characterization of a ParE-ParD toxin-antitoxin TA system (SO_A0087-A0088) on a megaplasmid of Shewanella oneidensis
Received date: 2018-02-28
Online published: 2018-12-24
Supported by
National Natural Science Foundation of China (31500025, 31500093)
Copyright
毒素-抗毒素系统在原核生物中分布十分广泛, 在细菌的生命活动中扮演了重要的角色, 如维持水平基因转移元件的稳定性以及应对环境胁迫等。然而目前对于来自生态环境菌株中的毒素-抗毒素系统的研究仍较少。文章鉴定了自然水体环境来源的菌株Shewanella oneidensis MR-1 携带的内源性大质粒上的一对ParE-ParD家族Ⅱ型毒素-抗毒素系统SO_A0088-A0087。毒素SO_A0088在大肠杆菌以及原宿主S. oneidensis 内均具有明显的细胞毒性, 并导致细胞分裂存在缺陷。抗毒素SO_A0087能够完全拮抗毒素的毒性。同时, 凝胶电泳迁移率实验(electrophoretic mobility shift assay, EMSA)证实抗毒素SO_A0087能够结合自身的启动子。另外, 文章还通过易错突变的方法找到了毒素SO_A0088的3个毒性关键位点。
关键词: 毒素-抗毒素系统; ParE-ParD; Shewanella oneidensis; 质粒
赵玄玉 , 施斐 , 王晓雪 . Shewanella oneidensis内源性大质粒上ParE-ParD家族毒素-抗毒素系统(SO_A0088-A0087)的鉴定[J]. 热带海洋学报, 2018 , 37(6) : 104 -111 . DOI: 10.11978/2018022
Toxin-antitoxin (TA) systems are prevalent in prokaryotes. They play important roles in cell physiological activities including maintenance of mobile genetic elements and mediation of stress responses. However, studies of TA systems in the bioelectrochemically and ecologically important strain Shewanella oneidensis are limited. In this study, we characterized SO_A0087-A0088 on the megaplasmid of Shewanella oneidensis MR-1 as a typical type Ⅱ TA system that belongs to ParE-ParD family, and the overexpression of toxin A0088 leads to filamentous growth. We found that the antitoxin SO_A0087 could bind to promoter of SO_A0087-A0088 in vitro, and three amino acid residues of toxin SO_A0088 were identified to be critical for toxicity by random mutagenesis.
Key words: toxin-antitoxin system; ParE-ParD; Shewanella oneidensis; plasmid
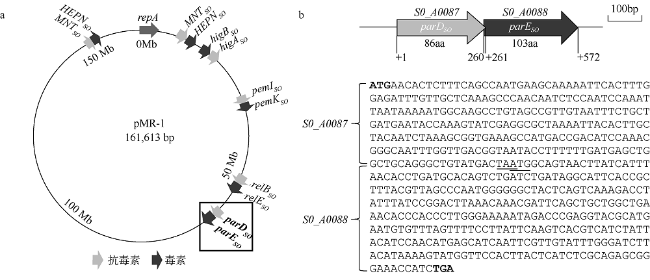
Fig. 1 Putative type II TA systems on megaplasmid of S. oneidensis MR-1 and characteristics of ParDE TA system. (a) Distribution of six putative type II TA systems on megaplasmid of S. oneidensis MR-1. |
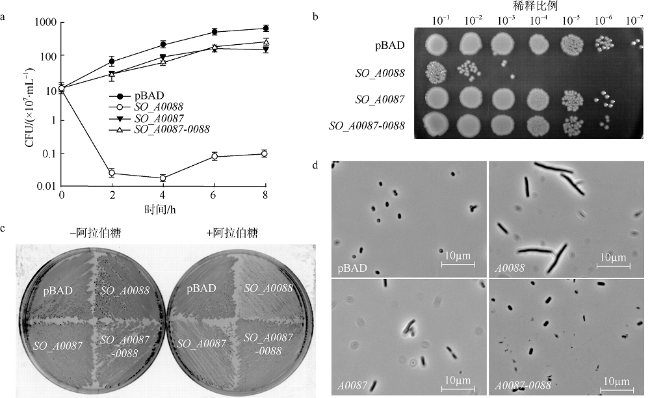
Fig. 2 Identification of SO_A0087-0088 as a TA system in S.oneidensis MR-1. |
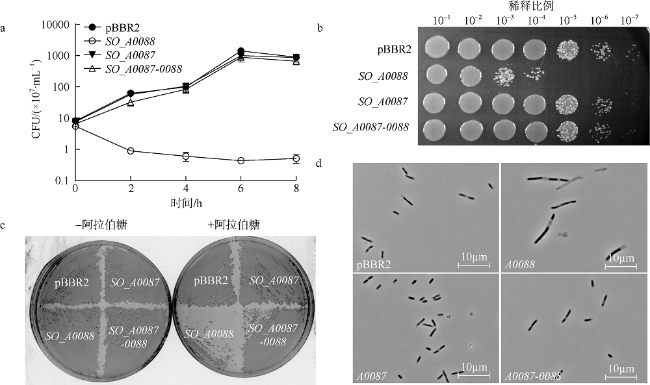
Fig. 3 Identification of SO_A0087_0088 as a TA system in E.coli BW25113. |
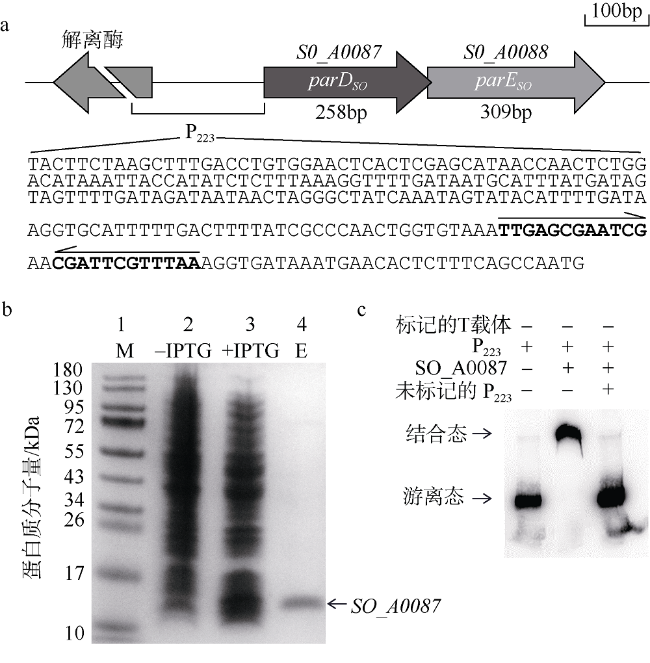
Fig. 4 Antitoxin SO_A0087 binds to promoter of SO_A0087- SO_A0088 in vitro. |
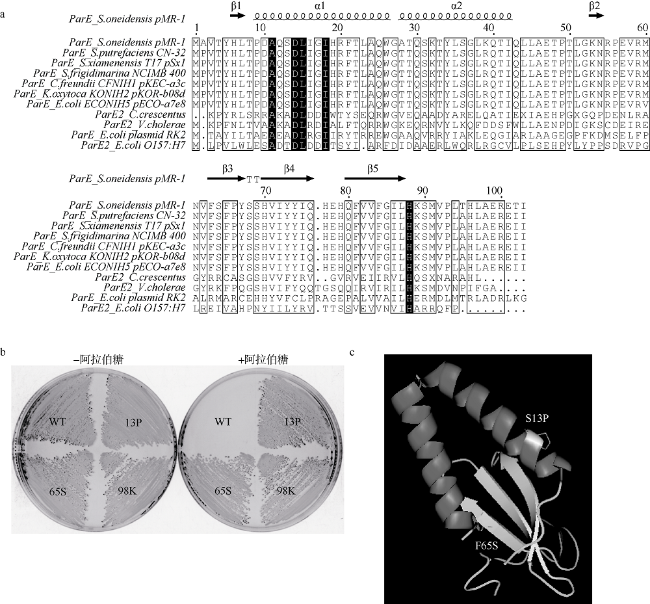
Fig. 5 ParE proteins alignments and mutation of active sites of SO_A0088. |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}