
海洋蛭弧菌DA5全基因组测序及序列分析
作者简介: 陆友云(1991—), 女, 湖南省娄底市人, 硕士研究生, 从事海洋微生物学研究。E-mail: youyunlu@126.com
收稿日期: 2018-02-06
网络出版日期: 2018-12-24
基金资助
国家自然科学基金项目(31372536);广东省科技计划项目(2015A020209160)
Whole-genome sequencing and analysis of marine Bdellovibrio-and-like organism DA5
Received date: 2018-02-06
Online published: 2018-12-24
Supported by
National Natural Science Foundation of China (31372536);Science and Technology Planning Project of Guangdong Province (2015A020209160)
Copyright
为深入挖掘和利用海洋蛭弧菌及其基因资源, 本研究利用Illumina HiSeq测序平台对一株属于嗜盐噬菌弧菌(Halobacteriovorax)的海洋蛭弧菌DA5进行了全基因组测序, 对基因组进行组装、基因预测和功能注释, 并与其他8株蛭弧菌进行了比较基因组分析。结果显示: DA5基因组大小为3.27Mb, GC含量为36.5%, 预测编码基因3175个。在DA5基因组中注释到303个基因具有直系同源蛋白簇分类, 与代谢通路相关基因1239个。比较基因组分析表明: DA5符合海洋蛭弧菌基因组的基本特征, 与其他8株蛭弧菌共有467个同源基因家族, 而DA5特有基因为266个。DA5中与细胞运动相关基因62个, 其中编码甲基受体趋化蛋白、鞭毛蛋白、鞭毛马达及其开关蛋白基因均与嗜盐噬菌弧菌BAL6_X的对应基因亲缘关系最近。对DA5全基因组序列的注释和功能分析, 为深入研究其捕食特性和作用机制, 并更有效地利用其防控海水养殖细菌病害提供了基础。
陆友云 , 薛明 , 李志桦 , 温崇庆 . 海洋蛭弧菌DA5全基因组测序及序列分析[J]. 热带海洋学报, 2018 , 37(6) : 112 -119 . DOI: 10.11978/2018016
To explore and utilize marine Bdellovibrio-and-like organisms (BALO) and their genes, the whole genome of Halobacteriovorax sp. DA5 was sequenced using the Illumina HiSeq platform. Then, fragment assembly, gene prediction and functional annotation were analyzed in comparison with those of other eight BALO strains. The results showed that the genome size of strain DA5 is 3.27 Mb with GC content of 36.5%, and encodes 3175 predicted genes, including 303 genes found in the Clusters of Orthologous Groups (COG) database and 1239 genes related to metabolic pathways. Comparative genomic analysis showed that DA5 has typical characteristics identified in genomes of the marine BALOs, and there were 467 homologous gene families common in genomes of DA5 and other eight BALOs, while 266 unique genes were found only in DA5 genome. Among all the 62 genes associated with cell motility of strain DA5, members such as methyl-accepting chemotaxis protein, putative flagellin, flagellar motor proteins, and flagellar motor switch proteins were most affiliated phylogenetically with those of Halobacteriovorax sp. BAL6_X. Therefore, annotation and functional analysis of the genome sequence of DA5 provide insight into its predatory property and interaction mechanism with host, and establish the basis for DA5 as biocontrol agent against harmful bacteria occurring in marine aquaculture.
Tab. 1 Gene distribution based on COG classification of Halobacteriovorax DA5表1 嗜盐噬菌弧菌DA5基因组COG功能分类 |
| 功能类别 | 基因数量 | 比例/% |
|---|---|---|
| 翻译、核糖体结构和生物合成 | 45 | 14.85 |
| 能量产生与转换 | 33 | 10.89 |
| 翻译后修饰、蛋白质周转和分子伴侣 | 30 | 9.90 |
| 氨基酸转运与代谢 | 21 | 6.93 |
| 无机离子转运与代谢 | 19 | 6.27 |
| 脂类转运与代谢 | 15 | 4.95 |
| 转录 | 15 | 4.95 |
| 一般功能预测 | 15 | 4.95 |
| 复制、重组和修复 | 14 | 4.62 |
| 核苷酸转运与代谢 | 11 | 3.63 |
| 细胞运动 | 11 | 3.63 |
| 碳水化合物的运输和代谢 | 10 | 3.30 |
| 信号转导机制 | 10 | 3.30 |
| 辅酶转运与代谢 | 8 | 2.64 |
| 细胞周期调控, 细胞分裂, 染色体分裂 | 5 | 1.65 |
| 细胞壁/膜/包体生物合成 | 5 | 1.65 |
| 防御机制 | 5 | 1.65 |
| 胞内运输、分泌和囊泡运输 | 3 | 0.99 |
| 次级代谢产物的生物合成、运输和分解代谢 | 2 | 0.66 |
| 细胞骨架 | 1 | 0.33 |
| 功能未知 | 25 | 8.25 |
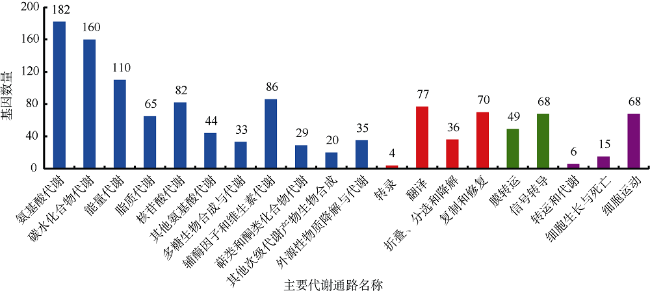
Fig. 1 The genetic metabolic pathways of Halobacteriovorax DA5图1 嗜盐噬菌弧菌DA5的基因代谢通路 |
Tab. 2 General features of genomes of Halobacteriovorax DA5 and reference strains表2 嗜盐嗜菌弧菌DA5与参考菌株基因组的基本特征 |
| 菌株 | 基因组大小/bp | 拼接水平 | GC含量/% | 预测基因总数 | 编码基因数量 | tRNA数量 |
|---|---|---|---|---|---|---|
| B. bacteriovorus HD100 | 3782950 | chromosome | 50.6 | 3597 | 3548 | 36 |
| H. marinus. SJ | 3435933 | chromosome | 36.7 | 3307 | 3254 | 36 |
| H. marinus. BE01 | 3393238 | chromosome | 36.7 | 3257 | 3201 | 36 |
| Halobacteriovorax sp. BSW11_IV | 3650096 | contig | 36.9 | 3491 | 3457 | 30 |
| Halobacteriovorax sp. SEQ25_V | 3450786 | contig | 36.0 | 3332 | 3292 | 32 |
| Halobacteriovorax sp. DB6_IX | 2969235 | contig | 37.6 | 3218 | 3192 | 23 |
| Halobacteriovorax sp. BAL6_X | 3233679 | contig | 36.5 | 3101 | 3065 | 31 |
| Halobacteriovorax sp. JY17 | 3470022 | contig | 36.1 | 3316 | 3263 | 33 |
| Halobacteriovorax sp. DA5 | 3272462 | contig | 36.5 | 3175 | 3125 | 31 |
注; 预测基因总数是指通过Glimmer 3.02软件预测到的基因数量; 编码基因数量是指蛋白质编码基因(CDS基因)数量。 |
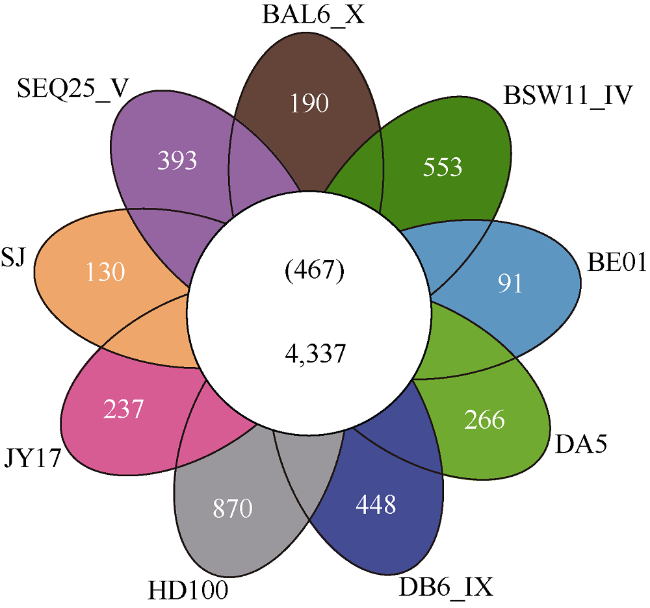
Fig. 2 Venn's diagram of homologous genes of Halobacteriovorax DA5 and other BALOs图2 嗜盐噬菌弧菌DA5与其他蛭弧菌的同源基因比较 |
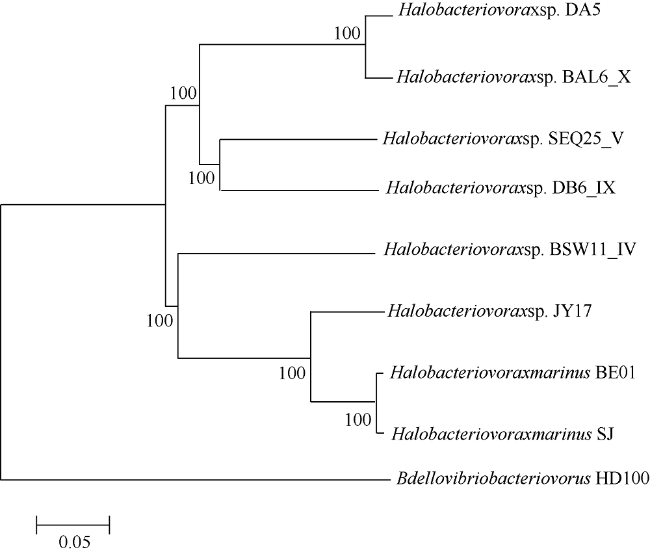
Fig. 3 Genome-wide phylogenetic tree of Halobacteriovorax DA5 and other BALOs. |
Tab. 3 Genes related to cell motility of Halobacteriovorax DA5表3 嗜盐噬菌弧菌DA5运动相关基因 |
| 基因名称 | 基因数量 | 编码蛋白名称 | 重叠群 |
|---|---|---|---|
| mcp | 7 | Methyl-accepting chemotaxis protein | contig1、contig3、contig7、contig9 |
| mcp | 1 | chemotaxis protein | contig7 |
| fliM/fliN/fliG | 3 | flagellar motor switch protein | contig3、contig3、contig9 |
| motB/motA | 2 | Flagellar motor protein | contig1 |
| motA | 2 | transporter, MotA/TolQ/ExbB proton channel family protein | contig1 |
| cheW | 1 | CheW-like protein | contig3 |
| cheY/cheA/cheW | 4 | putative chemotaxis protein | contig3、contig3、contig7 |
| cheY | 2 | response regulator receiver domain protein | contig1、contig8 |
| cheX | 1 | chemotaxis phosphatase | contig3 |
| cheB/cheR | 2 | protein-glutamate O-methyltransferase | contig3、contig9 |
| cheR | 1 | putative chemotaxis protein methyltransferase | contig7 |
| cheD | 1 | putative methylation of MCP proteins-related protein | contig7 |
| aer | 1 | hypothetical protein | contig7 |
| dppA | 1 | putative dipeptide ABC transporter, periplasmic dipeptide-binding protein DppA | contig11 |
| MYLPF | 1 | Regulation of actin cytoskeleton | contig36、contig37 |
| fliC | 5 | putative flagellin protein | contig8、contig1 |
| fliC | 1 | flagellin N-terminal domain protein | contig12 |
| fliP/fliQ/flhB/flhA | 4 | flagellar biosynthetic protein | contig3 |
| flhF | 1 | hypothetical protein | contig3 |
| fliR | 1 | bacterial export protein, family 1 | contig3 |
| flhG, fleN | 1 | ATPase MipZ | contig3 |
| flhG, fleN | 1 | CobQ/CobB/MinD/ParA nucleotide binding domain protein | contig3 |
| fliS | 1 | flagellar protein | contig8 |
| fliD/flgL/flgK | 3 | flagellar hook-associated protein | contig8 |
| flgI | 1 | flagellar P-ring protein | contig8 |
| flgH | 1 | flagellar L-ring protein | contig8 |
| fliF | 1 | flagellar M-ring protein | contig9 |
| flgG/flgF/flgB/flgC | 4 | flagellar basal-body rod protein | contig8、contig8、contig9、contig9 |
| fliE | 1 | flagellar hook-basal body complex protein | contig9 |
| fliH | 1 | flagellar assembly protein | contig9 |
| fliI | 1 | flagellum-specific ATP synthase | contig9 |
| fliJ | 1 | flagellar FliJ protein | contig9 |
| flgD | 1 | putative flagellar hook capping protein | contig9 |
| flgE | 1 | flagellar hook-basal body protein | contig9 |
| pilT | 1 | twitching motility protein | contig8 |
Tab. 4 Amino acid sequence homology comparison of the flagellin, flagella motor proteins, and flagella motor switch proteins表4 鞭毛蛋白、鞭毛马达及开关蛋白基因的氨基酸序列同源性比较 |
| 基因名称 | 比较菌株与序列相似性/% | ||
|---|---|---|---|
| DA5与SJ | DA5与BAL6_X | SJ与BAL6_X | |
| fliC1 | 67 | 97 | 68 |
| fliC2 | 64 | 95 | 65 |
| fliC3 | 92 | 100 | 92 |
| fliC4 | 77 | 96 | 77 |
| fliC5 | 79 | 98 | 78 |
| motB | 66 | 98 | 67 |
| motA | 75 | 99 | 74 |
| fliM | 86 | 99 | 86 |
| fliN | 82 | 100 | 82 |
| fliG | 93 | 100 | 93 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}