
模拟升温和营养盐加富对大亚湾浮游生物群落代谢的影响*
作者简介:谢福武(1992—), 男, 海南省万宁市人, 从事海洋生态学研究。E-mail: xfw1021900977@163.com
收稿日期: 2018-07-25
要求修回日期: 2018-09-10
网络出版日期: 2019-04-15
基金资助
国家重点研发计划(2017YFC0506302)
国家重点基础研究发展计划(2015CB452904)
国家自然科学基金(41276161)
Impact of simulated warming and nutrients input on plankton community metabolism in Daya Bay*
Received date: 2018-07-25
Request revised date: 2018-09-10
Online published: 2019-04-15
Supported by
National Key Research and Development Program of China (2017YFC0506302)
National Basic Research Program (2015CB452904)
National Natural Science Foundation of China (41276161)
Copyright
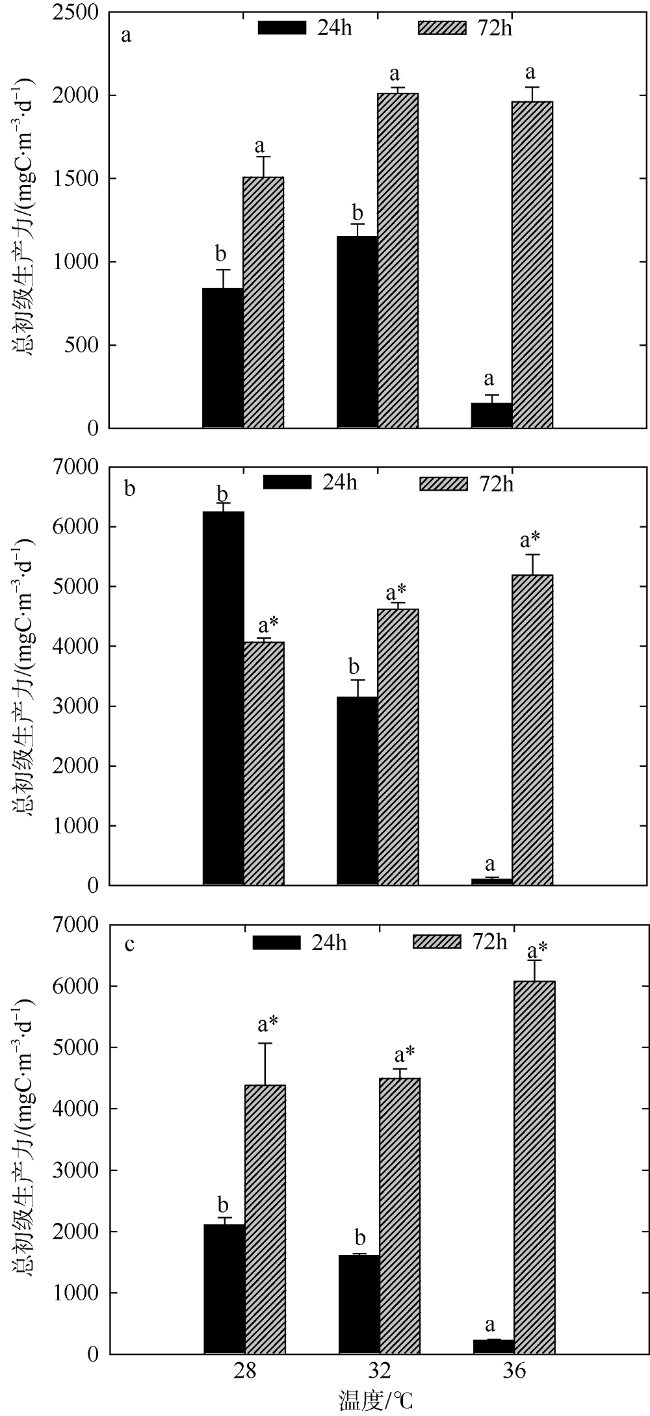
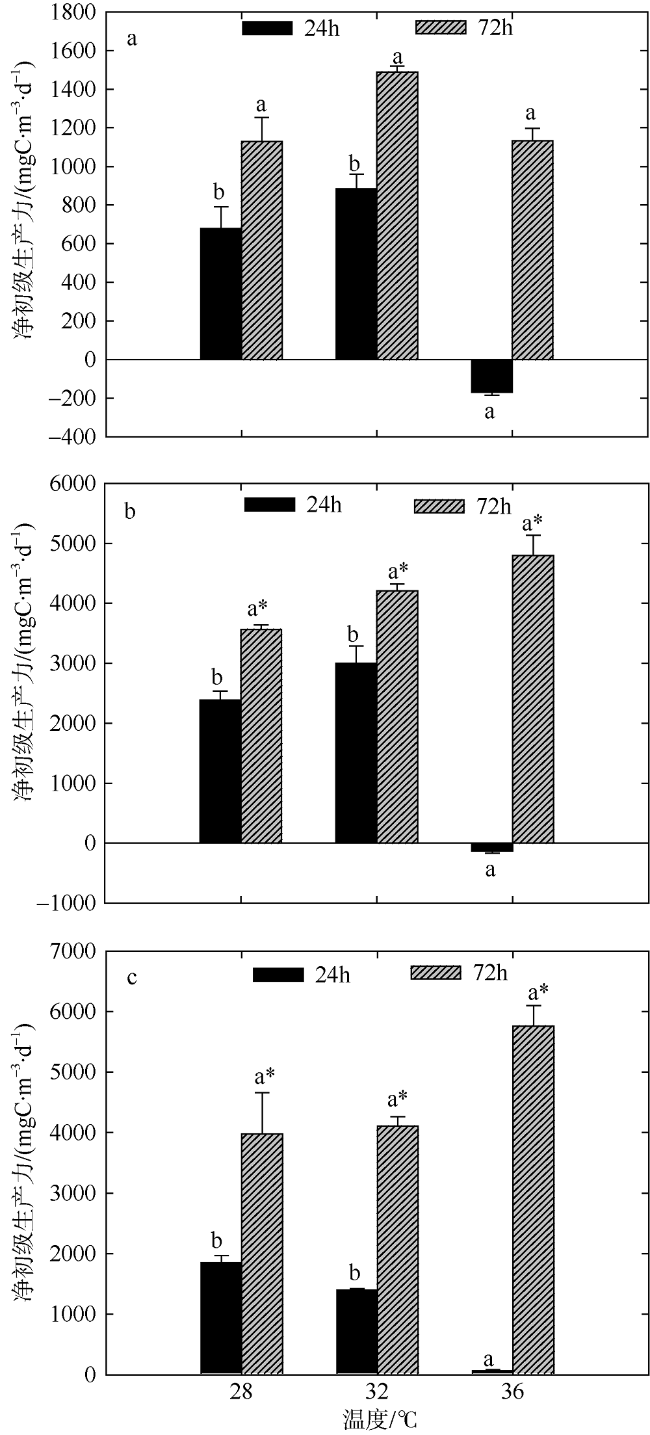
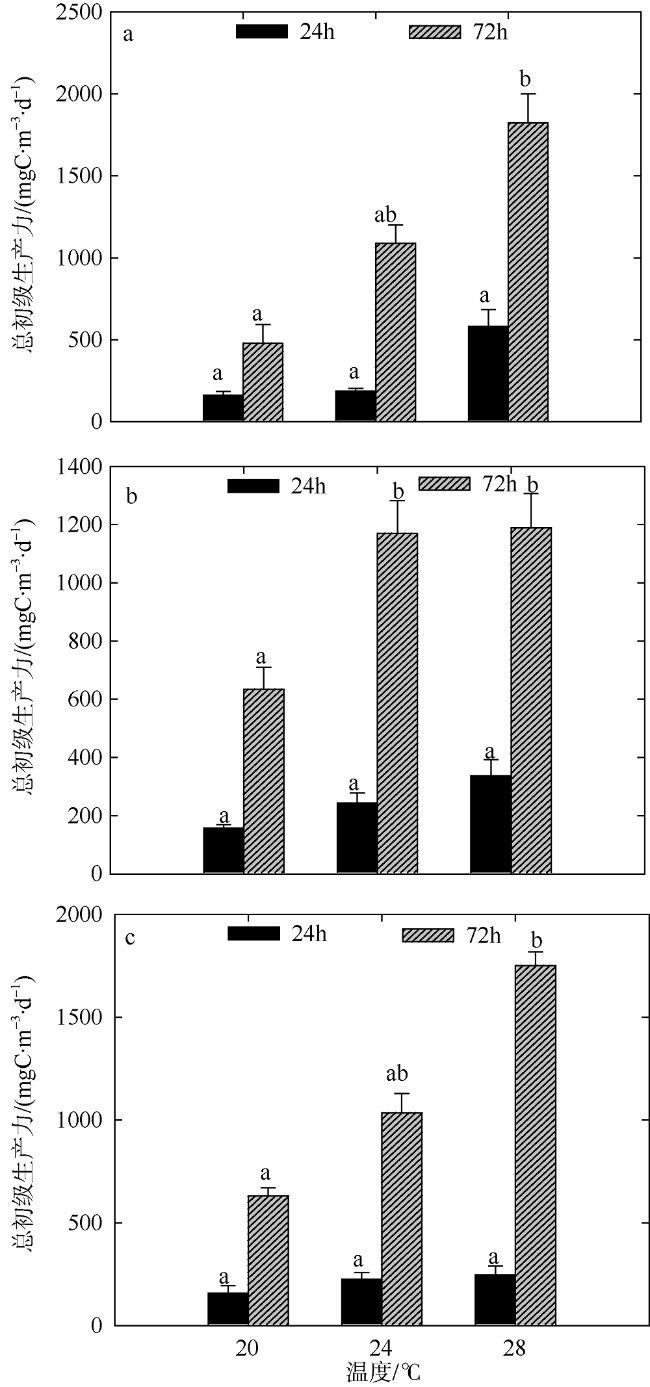
文章结合模拟升温和营养盐加富, 于2016年8月和2017年1月研究了夏季和冬季大亚湾海域浮游生物群落代谢特征, 以期深入了解浮游生物对升温和富营养化作用的响应机制。结果表明: 升温和营养盐输入均会影响浮游生物总初级生产力(GPP)和群落呼吸率(CR), 且GPP对环境的响应更敏感。夏季GPP除在培养前期(24h)极高温条件下受到显著抑制外, 营养盐输入的影响比温度的影响作用更显著; 而冬季GPP和CR受升温效应的影响显著(p<0.05)。总体看来, 升温效应和营养盐输入均能影响浮游生物群落碳代谢平衡, 导致海洋生态系统的稳定性降低, 进而影响海洋生物多样性以及渔业资源产出的数量和质量。
谢福武 , 宋星宇 , 谭烨辉 , 谭美婷 , 黄亚东 , 刘华雪 . 模拟升温和营养盐加富对大亚湾浮游生物群落代谢的影响*[J]. 热带海洋学报, 2019 , 38(2) : 48 -57 . DOI: 10.11978/2018075
Characters of plankton community metabolism of Daya Bay were studied using data collected in August 2016 and January 2017. The possible effects of nutrients input were discussed to understand the response mechanism of plankton to warming and eutrophication based on simulation experiments. The results indicated that both GPP (gross primary production) and CR (community respiration) were influenced by warming and nutrients input, and response of GPP to environment was seriously sensitive. The impact of GPP on nutrients input was more remarkably than that of temperature, except for being significantly inhibited in early cultured stage (24 h) with extremely high temperature conditions. In winter, both GPP and CR were significantly affected by the warming effect (p<0.05). Overall, both warming effect and nutrients input could affect carbon metabolism of plankton community which would lead to the declining of marine ecosystem stability, thereby affecting marine ecosystem stability and biodiversity and quality of marine ecological environment and fishery resource output.
Key words: plankton; community metabolism; nutrients input; Daya Bay
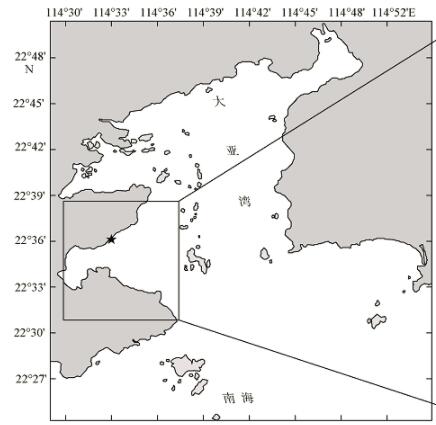
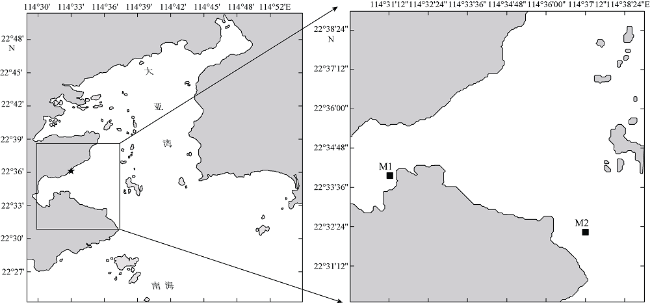
Fig. 1 Maps of sampling stations in Daya Bay图1 现场温度调查和模拟实验采样站位分布图 |
Tab. 1 Simulation experimental design in Daya Bay during summer and winter表1 大亚湾模拟实验设计 |
| 单因素模拟实验条件(M1) | 交互模拟实验(M2) | |
|---|---|---|
| 夏季 | 6个温度梯度(38℃、36℃、34℃、32℃、30℃、28℃) | 3个营养盐加富梯度*和3个温度梯度(36℃、32℃、28℃) |
| 冬季 | 6个温度梯度(28℃、26℃、24℃、22℃、20℃、18℃) | 3个营养盐加富梯度*和3个温度梯度(28℃、24℃、20℃) |
注: *营养盐加富梯度[低浓度组(LN): 不添加营养盐; 中浓度组(MN): 添加终浓度为10μmol·L-1的硝酸盐及0.6μmol·L-1的磷酸盐; 高浓度组(HN): 添加终浓度为100μmol·L-1的硝酸盐及6μmol·L-1的磷酸盐] |
Tab. 2 The environmental parameters of surface water in Daya Bay表2 夏冬两季大亚湾表层站位环境参数 |
| 季节 | 站位 | 温度/℃ | 盐度/‰ | pH | Chl-a/(μg·L-1) | DIP/(μmol·L-1) | DIN/(μmol·L-1) | DIN/DIP |
|---|---|---|---|---|---|---|---|---|
| 夏季 | M1 | 28.3 | 30.7 | 8.1 | 5.37 | 0.44 | 35.46 | 80.59 |
| M2 | 29.9 | 33.6 | 8.3 | 1.58 | 0.05 | 7.82 | 156.40 | |
| 冬季 | M1 | 18.4 | 29.1 | 8.0 | 0.63 | 0.51 | 12.50 | 24.51 |
| M2 | 17.9 | 29.9 | 8.0 | 1.43 | 0.30 | 10.62 | 35.40 |
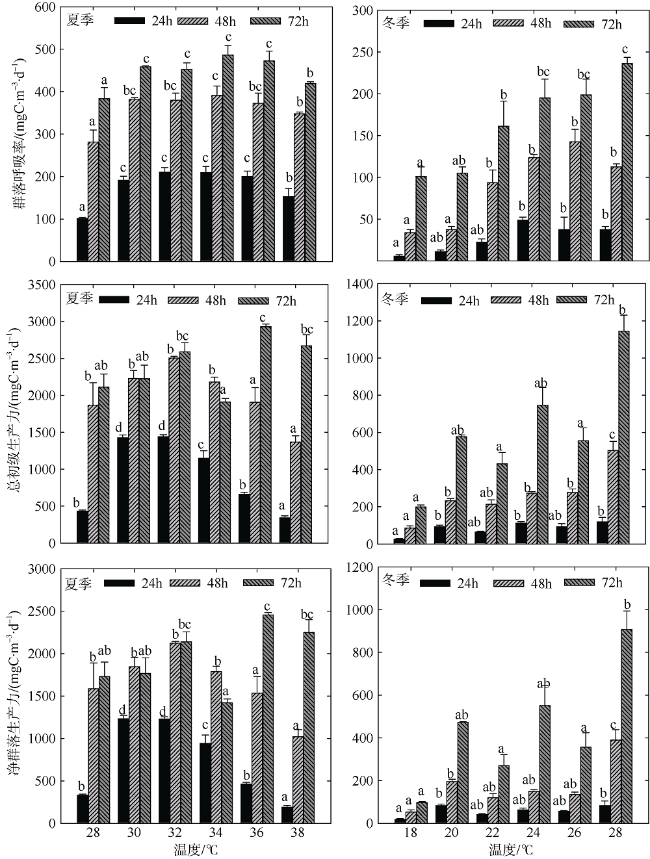
Fig. 2 Features of plankton community metabolism in simulated experiments图2 模拟升温实验浮游生物群落代谢特征 |
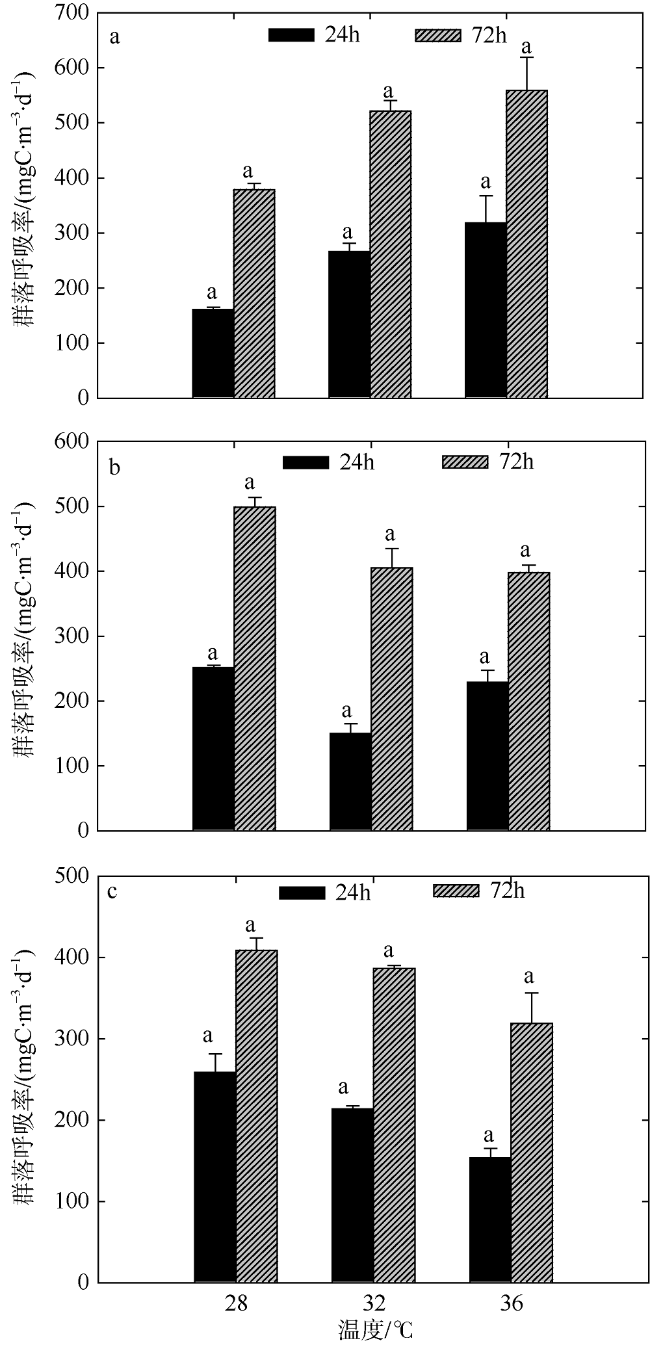
Fig. 3 Variation of plankton CR of two-factors simulated experiments in summer. Significant differences among the temperature groups are indicated by using different superscripts (p<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”图3 夏季双因素模拟实验浮游生物群落呼吸率的变化 |
Fig. 4 Variation of plankton GPP of two-factors simulated experiments in summer. Significant differences among the temperature groups are indicated by using different superscripts (p<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”图4 夏季双因素模拟实验浮游生物群落总初级生产力的变化 |
Fig. 5 Variation of plankton NCP of two-factors simulated experiments in summer. Significant differences among the temperature groups are indicated by using different superscripts (p<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”图5 夏季双因素模拟实验浮游生物群落净生产力的变化 |
Tab. 3 Ratio of GPP/CR of two-factors simulated experiments during summer and winter表3 TSE浮游生物总生产与总代谢的比值(GPP/CR) |
| 营养盐 | 夏季 | 冬季 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 28℃ | 32℃ | 36℃ | 均值 | 20℃ | 24℃ | 28℃ | 均值 | ||
| 24h | 低营养 | 5.21 | 4.32 | 0.47 | 3.33 | 5.38 | 2.78 | 1.76 | 3.31 |
| 中营养 | 10.50 | 21.00 | 0.44 | 10.65 | 2.63 | 2.60 | 2.37 | 2.53 | |
| 高营养 | 8.14 | 7.53 | 1.44 | 5.70 | 2.63 | 3.00 | 1.40 | 2.34 | |
| 均值 | 7.95 | 10.95 | 0.78 | 3.55 | 2.79 | 1.84 | |||
| 72h | 低营养 | 3.98 | 3.86 | 10.72 | 6.19 | 2.51 | 5.92 | 3.12 | 3.85 |
| 中营养 | 8.15 | 11.39 | 11.63 | 10.39 | 2.91 | 4.52 | 3.14 | 3.52 | |
| 高营养 | 10.72 | 13.18 | 19.07 | 14.32 | 2.95 | 4.00 | 4.41 | 3.79 | |
| 均值 | 7.62 | 9.48 | 13.81 | 2.79 | 4.81 | 3.56 | |||
Fig. 6 Variation of plankton CR of two-factors simulated experiments in winter. Significant differences among the temperature groups are indicated by using different superscripts (p<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”图6 冬季双因素模拟实验浮游生物群落CR值变化 |
Fig. 7 Variation of plankton GPP of two-factors simulated experiments in winter. Significant differences among the temperature groups are indicated by using different superscripts (p<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”图7 冬季双因素模拟实验浮游生物群落GPP值变化 |
Fig. 8 Variation of plankton NCP of two-factors simulated experiments in winter. Significant differences among the temperature groups are indicated by using different superscripts (p<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”图8 冬季双因素模拟实验浮游生物群落NCP值变化 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}