
基于28S rDNA的南海刺长腹剑水蚤(Oithona setigera)种群遗传多样性研究
|
季莹莹(1994—), 女, 江苏省南京市人, 硕士研究生, E-mail:15705117350@163.com |
Copy editor: 林强
收稿日期: 2018-10-25
要求修回日期: 2018-12-20
网络出版日期: 2019-06-17
基金资助
国家自然科学基金(41406188)
中国水产科学研究院南海水产研究所中央级公益性科研院所基本科研业务费专项资金(2017YB26、2016TS24)(2017YB26)
中国水产科学研究院南海水产研究所中央级公益性科研院所基本科研业务费专项资金(2017YB26、2016TS24)(2016TS24)
版权
Genetic structure of Oithona setigera from South China Sea based on 28S rDNA gene
Copy editor: LIN Qiang
Received date: 2018-10-25
Request revised date: 2018-12-20
Online published: 2019-06-17
Supported by
National Natural Science Foundation of China(41406188)
South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Scientific Research Funds for Central Non-profit Institutes(2017YB26)
South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Scientific Research Funds for Central Non-profit Institutes(2016TS24)
Copyright
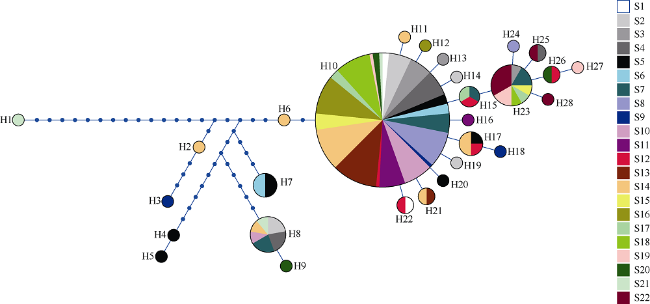
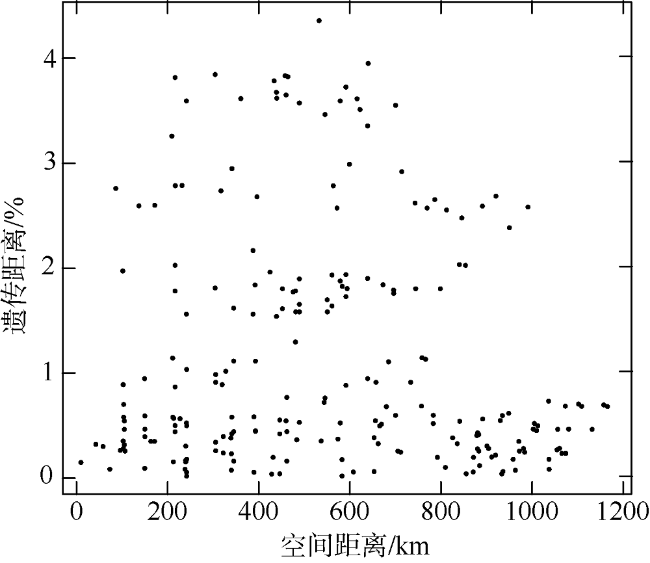
长腹剑水蚤属是海洋中小型浮游动物中最为丰富的类群之一, 在生物地理学与海洋生态学研究中均具有重要地位。本研究基于28S rDNA分析了南海长腹剑水蚤属中较为常见的刺长腹剑水蚤Oithona setigera的单倍型多样性和种群遗传结构。结果显示, 792bp长度的核苷酸片段中, 碱基G+C的平均含量为58.2%, 高于A+T含量(41.8%)。种群平均遗传距离ΦST为0.011。在22个种群共计186个个体中, 发现了28个单倍型, 其中单倍型H10在21个种群中均被发现, 最远距离超过1000km, 说明刺长腹剑水蚤可以实现远距离的扩散且受到南海海流影响。Mantel检验结果显示, 刺长腹剑水蚤种群遗传距离和地理距离无线性相关性(R=-0.04615, P=0.678); RDA变差分解结果显示, 空间变量全模型对种群遗传结构的解释率为53.3%, 结合种群平均遗传距离ΦST为0.011, 我们判断目前观测到的刺长腹剑水蚤的种群遗传结构可能由历史上种群扩展带来的拓殖隔离造成。
季莹莹 , 徐磊 , 黎红 , 王亮根 , 杜飞雁 . 基于28S rDNA的南海刺长腹剑水蚤(Oithona setigera)种群遗传多样性研究[J]. 热带海洋学报, 2019 , 38(3) : 89 -97 . DOI: 10.11978/2018112
Oithona is one of the most abundant species of small and medium zooplankton in the ocean, and plays an important role in marine biogeographic genetics and ecology research. In this study, we employed 28S rDNA to analyze the population genetic structure and haplotype pattern of the most common species (Oithona setigera) in the South China Sea. A dataset of 792 bp in length sequences was obtained. The average contents of G+C (58.2%) were significantly higher than those of A+T (41.8%) in the fragment. A total of 28 haplotypes were defined from 186 individuals, of which the dominant haplotype H10 was found in 21 populations. The largest distance between two sampling sites harboring this haplotype is more than 1000 km, indicating Oithona setigera can achieve long distance disperse and be affected by ocean currents. The Mantel test showed that there was no linear correlation between the genetic distance and geographical distance (R= -0.04615, P=0.678); the RDA (redundancy analysis) results indicated space factor significantly affect the population genetic structure rather than environmental factors. The population genetic structure of Oithona setigera may be caused by colonization events followed by demographic expansions.
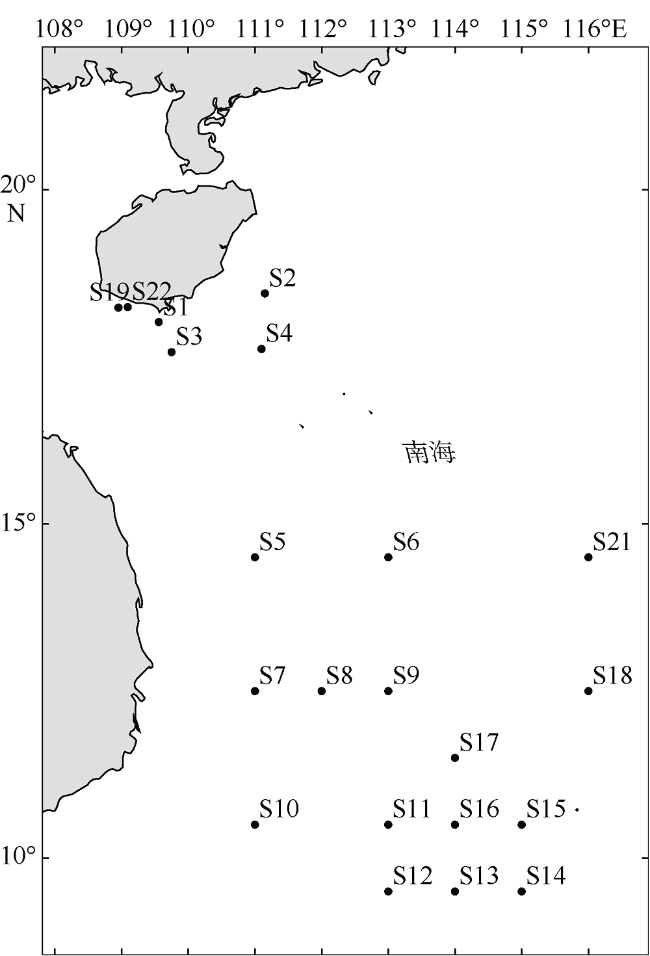
表1 刺长腹剑水蚤(Oithona setigera) 种群采集站点及其基本信息Tab. 1 Information of sample sites of Oithona setigera |
| 站位 | 纬度 | 经度 | 平均水温/℃ | 风速/(m·s-1) | 盐度/‰ | 叶绿素a/(mg·m-3) | 样本 数量 | 单倍型 数量 | 单倍型 |
|---|---|---|---|---|---|---|---|---|---|
| S1 | 18°15′36″N | 109°33'36"E | 24.49 | 8.19 | NaN | 0.23 | 3 | 2 | H10、H22 |
| S2 | 18°26'59N | 111°9'E | 24.44 | 8.95 | NaN | 0.12 | 11 | 4 | H8、H10、H14、H19 |
| S3 | 17°34'12"N | 109°45'E | 24.60 | 8.71 | NaN | 0.13 | 9 | 3 | H8、H10、H13 |
| S4 | 17°37'12"N | 111°5'59"E | 24.71 | 9.35 | 33.24 | 0.11 | 11 | 2 | H10、H25 |
| S5 | 14°30'N | 111°E | 26.54 | 10.78 | 33.36 | 0.08 | 9 | 5 | H4、H5、H7、H10、H20 |
| S6 | 14°30'N | 113°E | 26.93 | 11.04 | 33.25 | 0.08 | 5 | 2 | H7、H10 |
| S7 | 12°30'N | 111°E | 26.84 | 10.73 | 34.12 | 0.11 | 11 | 4 | H8、H10、H15、H13 |
| S8 | 12°30'N | 112°E | 27.09 | 10.39 | 33.67 | 0.10 | 12 | 1 | H10、H24 |
| S9 | 12°30'N | 113°E | 27.23 | 10.13 | 33.50 | 0.08 | 3 | 2 | H10、H18 |
| S10 | 10°30'N | 111°E | 27.20 | 5.05 | 33.14 | 0.08 | 10 | 4 | H8、H10、H23、H27 |
| S11 | 10°30'N | 113°E | 27.36 | 4.35 | 33.53 | 0.10 | 9 | 2 | H10、H16 |
| S12 | 9°30'N | 113°E | 27.39 | 3.43 | 33.57 | 0.09 | 5 | 5 | H10、H22、H17、H15、H26 |
| S13 | 9°30'N | 114°E | 27.92 | 3.57 | 33.21 | 0.09 | 15 | 2 | H10、H21 |
| S14 | 9°30'N | 115°E | 28.13 | 5.88 | 33.36 | 0.11 | 20 | 6 | H2、H6、H8、H10、H11、H17、H21 |
| S15 | 10°30'N | 115°E | 27.97 | 7.62 | 33.32 | 0.10 | 6 | 2 | H10、H23 |
| S16 | 10°30'N | 114°E | 27.77 | 6.48 | 33.06 | 0.09 | 12 | 2 | H10、H12 |
| S17 | 11°30'N | 114°E | 27.57 | 9.17 | 33.25 | 0.11 | 6 | 4 | H1、H10、H15、H23 |
| S18 | 12°30'N | 116°E | 27.83 | 10.00 | 33.46 | 0.08 | 12 | 2 | H10、H23 |
| S19 | 18°14'23"N | 108°57'E | 25.06 | 7.77 | NaN | 0.98 | 4 | 1 | H10 |
| S20 | 14°30'N | 117°E | 27.50 | 10.60 | 33.42 | 0.10 | 4 | 2 | H9、H10、H26 |
| S21 | 14°30'N | 116°E | 27.55 | 10.90 | 33.36 | 0.09 | 2 | 2 | H8、H10 |
| S22 | 18°15'N | 109°5'59"E | 24.76 | 7.89 | NaN | 0.59 | 7 | 3 | H23、H25、H28 |
注: NaN为无数据 |
表2 部分站位之间刺长腹剑水蚤(Oithona setigera)的遗传距离(%)Tab. 2 Genetic distance (%) of Oithona setigera between two sampling sites |
| S8 | S9 | S10 | S11 | S12 | S13 | S14 | S15 | S16 | S17 | S18 | S19 | S20 | S21 | S22 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S9 | 0.89 | ||||||||||||||
| S10 | 0.18 | 0.99 | |||||||||||||
| S11 | 0.02 | 0.87 | 0.16 | ||||||||||||
| S12 | 0.16 | 1.02 | 0.30 | 0.16 | |||||||||||
| S13 | 0.45 | 1.12 | 0.58 | 0.43 | 0.58 | ||||||||||
| S14 | 0.45 | 1.12 | 0.56 | 0.42 | 0.57 | 0.70 | |||||||||
| S15 | 0.06 | 0.91 | 0.20 | 0.04 | 0.17 | 0.47 | 0.47 | ||||||||
| S16 | 0.26 | 1.04 | 0.40 | 0.24 | 0.40 | 0.55 | 0.59 | 0.28 | |||||||
| S17 | 0.09 | 0.95 | 0.23 | 0.08 | 0.18 | 0.50 | 0.50 | 0.09 | 0.32 | ||||||
| S18 | 0.04 | 0.89 | 0.18 | 0.02 | 0.16 | 0.45 | 0.45 | 0.06 | 0.26 | 0.09 | |||||
| S19 | 0.25 | 1.13 | 0.42 | 0.26 | 0.29 | 0.69 | 0.68 | 0.24 | 0.50 | 0.22 | 0.25 | ||||
| S20 | 0.53 | 1.30 | 0.60 | 0.52 | 0.60 | 0.95 | 0.89 | 0.53 | 0.76 | 0.55 | 0.53 | 0.62 | |||
| S21 | 1.78 | 2.17 | 1.79 | 1.76 | 1.90 | 1.94 | 1.94 | 1.81 | 1.90 | 1.84 | 1.78 | 2.03 | 1.98 | ||
| S22 | 0.26 | 1.15 | 0.43 | 0.28 | 0.27 | 0.71 | 0.70 | 0.24 | 0.52 | 0.20 | 0.26 | 0.15 | 0.60 | 2.04 |
注: 由于数据量大, 故只展示部分显著性较强的数据 |
表3 部分站位之间刺长腹剑水蚤(Oithona setigera)的空间距离(km)Tab. 3 Spatial distance (km) of Oithona setigera between two sampling sites |
| S8 | S9 | S10 | S11 | S12 | S13 | S14 | S15 | S16 | S17 | S18 | S19 | S20 | S21 | S22 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 660 | 740 | 878 | 940 | 1043 | 1087 | 1138 | 1044 | 987 | 891 | 943 | 65 | 898 | 805 | 49 |
| S2 | 669 | 691 | 885 | 907 | 1016 | 1043 | 1079 | 977 | 937 | 832 | 842 | 234 | 764 | 679 | 218 |
| S3 | 614 | 664 | 799 | 862 | 965 | 1009 | 1063 | 970 | 911 | 816 | 877 | 113 | 847 | 751 | 102 |
| S4 | 578 | 605 | 793 | 819 | 927 | 957 | 997 | 897 | 852 | 749 | 776 | 238 | 720 | 629 | 223 |
| S5 | 248 | 311 | 445 | 495 | 598 | 645 | 707 | 622 | 552 | 466 | 585 | 470 | 647 | 539 | 464 |
| S6 | 248 | 223 | 495 | 445 | 557 | 567 | 598 | 495 | 458 | 351 | 394 | 600 | 431 | 323 | 590 |
| S7 | 109 | 217 | 223 | 312 | 399 | 468 | 550 | 490 | 396 | 345 | 543 | 676 | 687 | 585 | 672 |
| S8 | 109 | 248 | 248 | 351 | 399 | 468 | 396 | 312 | 245 | 435 | 718 | 585 | 487 | 712 | |
| S9 | 312 | 223 | 334 | 351 | 399 | 312 | 248 | 156 | 326 | 773 | 487 | 394 | 765 | ||
| S10 | 219 | 246 | 347 | 452 | 438 | 328 | 346 | 589 | 889 | 789 | 702 | 887 | |||
| S11 | 246 | 347 | 452 | 452 | 328 | 346 | 589 | 889 | 789 | 702 | 887 | ||||
| S12 | 110 | 220 | 246 | 156 | 248 | 468 | 1067 | 707 | 645 | 1061 | |||||
| S13 | 110 | 156 | 111 | 223 | 399 | 1115 | 645 | 598 | 1108 | ||||||
| S14 | 111 | 156 | 248 | 351 | 1172 | 598 | 567 | 1164 | |||||||
| S15 | 109 | 156 | 248 | 1080 | 495 | 458 | 1072 | ||||||||
| S16 | 111 | 312 | 1019 | 552 | 495 | 1011 | |||||||||
| S17 | 245 | 926 | 466 | 398 | 918 | ||||||||||
| S18 | 990 | 248 | 223 | 979 | |||||||||||
| S19 | 955 | 860 | 16 | ||||||||||||
| S20 | 108 | 941 | |||||||||||||
| S21 | 847 |
注: 由于数据量大, 故只展示部分显著性较强的数据 |
表4 空间变量与环境变量的RDA变差分解结果Tab. 4 The redundancy analysis results of spatial variables and environmental variables |
| RDA | R2 | R2adj | P | |
|---|---|---|---|---|
| S (空间变量) | Global model(全模型) | 0.759 | 0.638 | 0.003 |
| dbMEM2 | 0.281 | 0.015 | ||
| dbMEM4 | 0.190 | 0.010 | ||
| dbMEM6 | 0.120 | 0.025 | ||
| dbMEM7 | 0.099 | 0.019 | ||
| E(环境变量) | Global model(全模型) | 0.304 | 0.141 | 0.05 |
| FS(前向选择) | 0.181 | 0.094 | 0.03 | |
| Wind speed(风速) | 0.170 | 0.048 | ||
| S+E(环境与空间变量) | 0.722 | 0.611 | 0.002 | |
| S|E(单纯空间变量) | 0.557 | 0.533 | 0.003 | |
| E|S(单纯环境变量) | 0.165 | 0.077 | NS | |
| Shard(环境与空间变量共享) | 0.084 | |||
| Residuals(残差) | 0.389 |
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
杜飞雁, 王亮根, 王雪辉 , 等, 2016. 南沙群岛海域长腹剑水蚤(Oithona spp.)的种类组成、数量分布及其与环境因子的关系[J]. 海洋与湖沼, 47(6):1176-1184.
|
| [2] |
范启, 何舜平 , 2014. 长江流域䱗种群遗传多样性和遗传结构分析[J]. 水生生物学报, 38(4):627-635.
|
| [3] |
黄琦, 徐少林, 徐磊 , 等, 2017. 广东流溪河水库盔型溞(Daphnia galeata)休眠种群与现生种群的单倍型多样性和遗传分化[J]. 湖泊科学, 29(5):1209-1216.
|
| [4] |
李纯厚, 贾晓平, 蔡文贵 , 2004. 南海北部浮游动物多样性研究[J]. 中国水产科学, 11(2):139-146.
|
| [5] |
林元烧 , 2005. 中华哲水蚤种群遗传学研究[D]. 厦门: 厦门大学.
|
| [6] |
刘光兴, 林坚 , 2007. 遗传标记技术在海洋桡足类生物多样性和系统发生研究中的应用[J]. 中国海洋大学学报, 37(1):33-37.
|
| [7] |
曲若竹, 侯林, 吕红丽 , 等, 2004. 群体遗传结构中的基因流[J]. 遗传, 26(3):377-382.
|
| [8] |
孙柔鑫, 王彦国, 连光山 , 等, 2014. 海南岛西北沿岸海域浮游桡足类的分布及群落特征[J]. 生物多样性, 22(3):320-328.
|
| [9] |
田志富 , 2012. 基于RDA的白洋淀浮游植物群落结构动态特征分析[D]. 保定: 河北大学.
|
| [10] |
王敏晓 , 2010. 分子标记在中国近海浮游桡足类研究中的应用 [D]. 青岛:中国科学院研究生院(海洋研究所).
|
| [11] |
王兴霞, 徐磊, 王亮根 , 等, 2018. 基于COI基因序列的长腹剑水蚤系统进化关系[J]. 海洋学报, 40(6):92-103.
|
| [12] |
徐兆礼 , 2006. 中国海洋浮游动物研究的新进展[J]. 厦门大学学报(自然科学版), 45(S2):16-23.
|
| [13] |
薛亚东, 李丽, 吴巩胜 , 等, 2011. 景观遗传学: 概念与方法[J]. 生态学报, 31(6):1756-1762.
|
| [14] |
杨海军, 刘秦玉 , 1998. 南海海洋环流研究综述[J]. 地球科学进展, 13(4):364-368.
|
| [15] |
张才学, 龚玉艳, 王学锋 , 等, 2011. 湛江港湾浮游桡足类群落结构的季节变化和影响因素[J]. 生态学报, 31(23):7086-7096.
|
| [16] |
张武昌, 赵楠, 陶振铖 , 等, 2010. 中国海浮游桡足类图谱[M]. 北京: 科学出版社: 800.
|
| [17] |
赵静, 孙洋, 谭永安 , 等, 2014. 基于COI及28S rDNA序列分析的扶桑绵粉蚧地理科群的遗传分化研究[J]. 棉花学报, 26(2):130-137.
|
| [18] |
朱伟军, 孙照渤, 齐卫宁 , 1997. 南海季风爆发及其环流特征[J]. 南京气象学院学报, 20(4):440-446.
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}