
大亚湾大型海藻丛的大角玻璃钩虾种群分布特征
|
王卉(1985—), 女, 河北省石家庄市人, 硕士, 从事海洋环境生态学研究。E-mail: |
Copy editor: 孙淑杰
收稿日期: 2018-11-14
要求修回日期: 2019-01-04
网络出版日期: 2019-07-21
基金资助
国家重点研发计划项目(2017YFB0903703); 广东省自然科学基金项目 (2018A030313136); 广州市科技计划项目(201707010163)()
版权
The population distribution of Hyale grandicornis in macroalgae canopies of Daya Bay
Copy editor: SUN Shu-jie
Received date: 2018-11-14
Request revised date: 2019-01-04
Online published: 2019-07-21
Supported by
National Key Research and Development Program (2017YFB0903703); Natural Science Foundation of Guangdong Province (2018A030313136); Guangzhou Science and Technology Planning Project (201707010163)()
Copyright
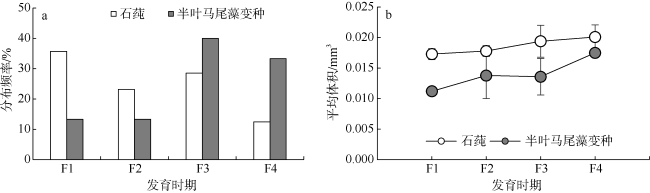
大角玻璃钩虾Hyale grandicornis是我国沿海潮间带常见的藻栖甲壳动物, 但目前关于其与栖息海藻的作用关系极少报道。2016年2—5月在大亚湾岩礁潮间带采集了石莼Ulva lactuca和半叶马尾藻变种Sargassum hemiphyllum var. chinense海藻, 结果显示大角玻璃钩虾在两种海藻丛中均有分布, 其中在石莼藻丛中分布的钩虾个体数量、平均体长和平均体重均大于在半叶马尾藻藻丛中。在半叶马尾藻藻丛中, 大角玻璃钩虾种群的性比、抱卵雌性比例和幼体比例相对较高, 而性成熟体长、抱卵雌性体长和卵粒体积则相对较小。研究表明, 大角玻璃钩虾在两种海藻中的种群分布与繁殖特征存在着差异, 可能受到栖息海藻形态结构及其生境等因素的影响。
王卉 , 李恒翔 , 李路 , 严岩 . 大亚湾大型海藻丛的大角玻璃钩虾种群分布特征[J]. 热带海洋学报, 2019 , 38(4) : 52 -58 . DOI: 10.11978/2018119
The gammarid Hyale grandicornis is a common crustacean species in macroalgae along the coastline of China, but the correlation between H. grandicornis and macroalgae is not well known. Two dominant macroalgae species found on coastal rocks of Daya Bay, the green algae Ulva lactuca and the brown algae Sargassum hemiphyllum var. chinense, were used in the experiments. H. grandicornis samples were found in the bush of two macroalgae canopies. The individual number, body size and weight of gammarid H. grandicornis population were higher in the bush of U. lactuca than in S. hemiphyllum var. chinense. The sex ratio and the proportion of females with eggs and the amount of juvenile of gammarid in bush of S. hemiphyllum var. chinense were larger than those in U. lactuca, whereas the body sizes of adults and females with eggs and the volume of eggs were smaller in gammarid from S. hemiphyllum var. chinense. Results demonstrate that the population distribution and production of gammarid H. grandicornis were different in the two macroalgae, which may be affected by the algal phenotype and habitat preference.
Key words: Daya Bay; gammarid; macroalgae; population distribution; production
表1 两种大型海藻丛的大角玻璃钩虾种群分布特征Tab. 1 The population distribution of Hyale grandicornis in two marine macroalgae |
| 大角玻璃钩虾 | 石莼藻丛中分布 | 半叶马尾藻变种藻丛中分布 |
|---|---|---|
| 个体总数 | 566 | 117 |
| 雌雄性比 | 1.10 | 1.46 |
| 幼体比例/% | 3.5 | 13.7 |
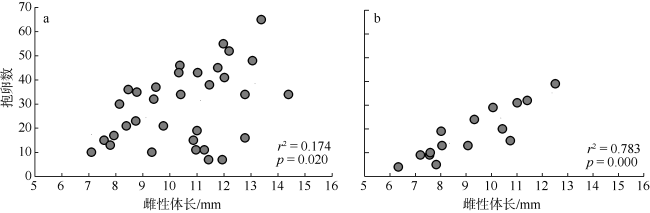
| 雌性抱卵率/% | 19.6 | 25.0 |
| 卵粒平均数 | 34.2±1.4 | 18.1±2.8 |
| 抱卵卵粒数 | 7~65 | 4~39 |
表2 两种大型海藻丛的大角玻璃钩虾种群体长分布Tab. 2 The body sizes of Hyale grandicornis in two marine macroalgae |
| 石莼藻丛 | 半叶马尾藻变种藻丛 | ||||
|---|---|---|---|---|---|
| 体长范围/mm | 平均体长±标准误差/mm | 体长范围/mm | 平均体长±标准误差/mm | ||
| 成熟雄性 | 7.06~18.40 | 11.56±0.10a | 5.46~16.31 | 10.04±0.40a | |
| 成熟雌性 | 5.00~14.73 | 9.58±0.13b | 5.09~13.22 | 8.33±0.28b | |
| 抱卵雌性 | 7.09~14.39 | 10.71±0.22ab | 6.33~12.51 | 9.14±0.47ab | |
| 幼体 | 1.63~4.96 | 4.19±0.14c | 1.50~4.86 | 3.53±0.27c | |
注: 凡标记相同小写字母的表示差异不显著(p>0.05), 不同小写字母的表示差异显著(p<0.05)。 |
表3 两种大型海藻丛的大角玻璃钩虾种群体重分布Tab. 3 The body weight of Hyale grandicornis in two marine macroalgae |
| 石莼藻丛 | 半叶马尾藻变种藻丛 | |||||
|---|---|---|---|---|---|---|
| 体重范围/mg | 平均体重±标准误差/mg | 体重范围/mg | 平均体重±标准误差/mg | |||
| 成熟雄性 | 5.4~26.0 | 12.3±0.6a | 2.8~24.8 | 11.5±0.9a | ||
| 成熟雌性 | 2.0~29.4 | 10.9±0.7b | 2.3~20.7 | 8.7±0.5b | ||
| 抱卵雌性 | 11.3~29.4 | 13.6±0.11a | 10.5~20.7 | 15.3±1.0c | ||
| 幼体 | 1.1~3.5 | 2.2±0.2c | 0.1~1.4 | 0.6±0.1d | ||
注: 凡标记相同小写字母的表示差异不显著(p>0.05), 不同小写字母的表示差异显著(p<0.05)。 |
| [1] |
任先秋 , 2006. 中国动物志无脊椎动物第四十一卷甲壳动物亚门端足目钩虾亚目(一)[M]. 北京: 科学出版社.
|
| [2] |
王友绍 , 2014. 大亚湾生态环境与生物资源[M]. 北京: 科学出版社.
|
| [3] |
郑新庆, 黄凌风, 王蕾 , 等, 2011. 筼筜湖大型海藻群落的几种藻栖端足类的种群动态研究[J]. 厦门大学学报(自然科学版), 50(5):928-933.
|
| [4] |
郑新庆, 黄凌风, 李元超 , 等, 2013. 啃食性端足类强壮藻钩虾对筼筜湖三种大型海藻的摄食选择性[J]. 生态学报, 33(22):7166-7172.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}