
防城河口湾鱼类群落结构及其与环境因子关系研究
|
何思璇(1997—), 女, 广西省东兴市人。主要从事生态学研究。E-mail: hesixuan2018@126.com |
Copy editor: 殷波
收稿日期: 2019-01-01
要求修回日期: 2019-04-28
网络出版日期: 2019-10-09
基金资助
国家重点研发计划重点专项(2017YFC0506100)
国家海洋局专项(YLFCJ20164006-F)
广西海洋局研究专项(GXHYJ100)
广西科技计划项目(AA17129002)
版权
Study on fish community structure and its relationship with environmental factors in Fangchenghe Estuary of Guangxi, China
Copy editor: YIN Bo
Received date: 2019-01-01
Request revised date: 2019-04-28
Online published: 2019-10-09
Supported by
National Key Research and Development Program of China(2017YFC0506100)
Science and Technology Project of State Oceanic Administration(YLFCJ20164006-F)
Science and Technology Project of Guangxi Oceanic Administration(GXHYJ100)
Guangxi Science and Technology Project(AA17129002)
Copyright
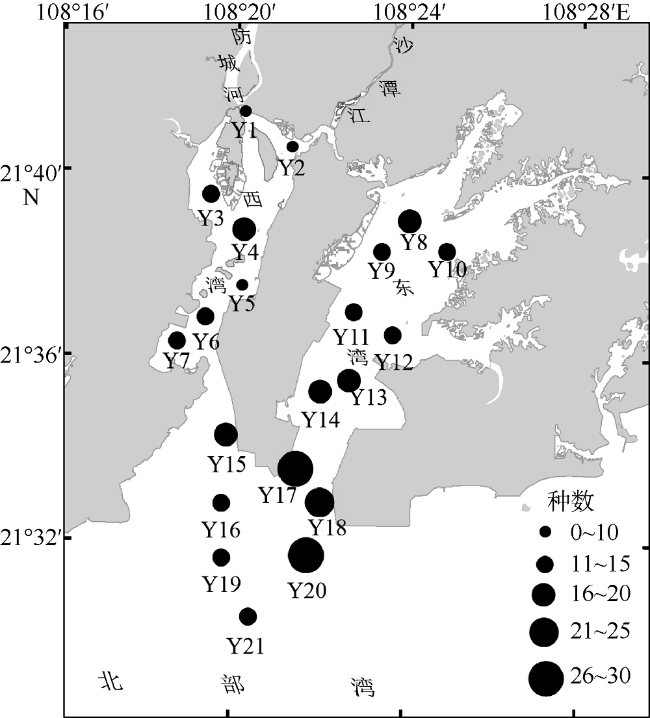
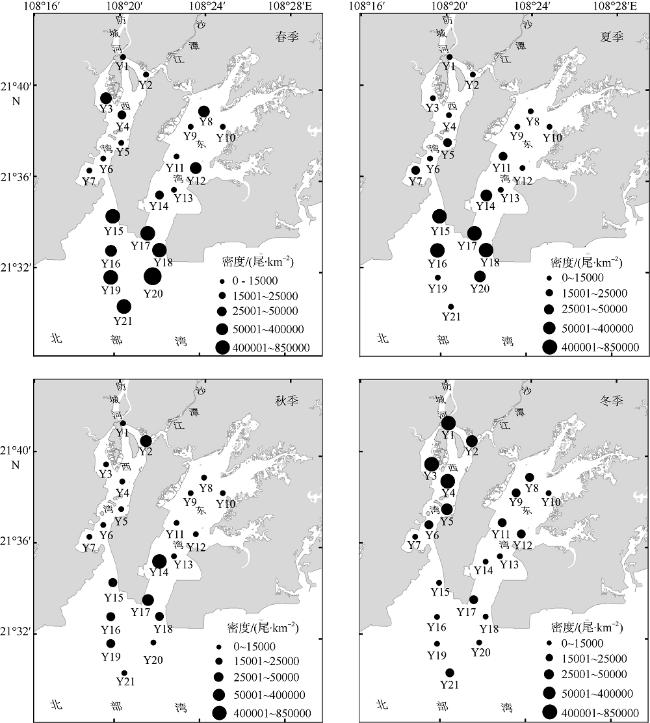
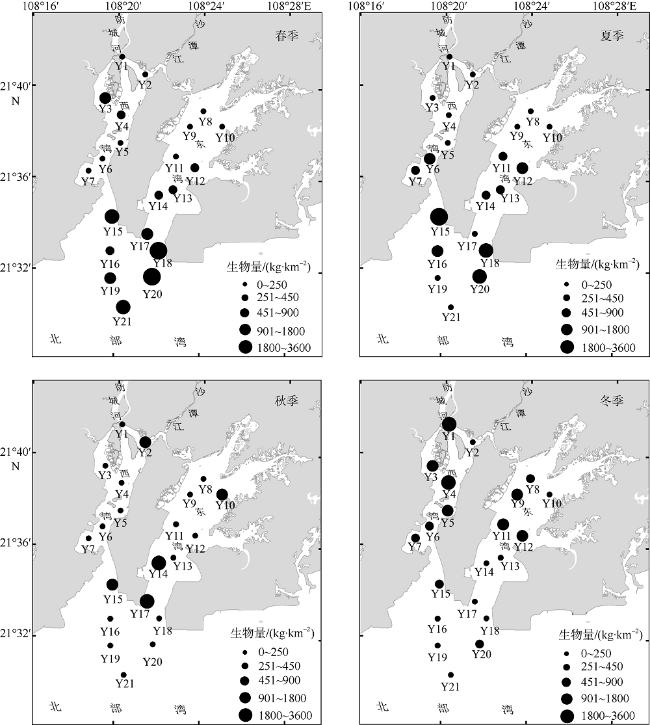
文章根据2016年5月、8月、11月和2017年2月在防城河口湾海域进行的渔业资源和环境调查数据, 研究了河口湾鱼类群落结构及其空间和季节变化, 分析了鱼类群落结构与主要环境因子的关系。结果表明: 全年调查共渔获鱼类95种, 隶属于2纲12目37科69属, 适温性以暖水性鱼类为主, 生态类型以底层和近底层鱼类为主。其中, 春季60种, 夏季59种, 秋季54种, 冬季46种, 夏—秋季间种类相似性指数最高。全年优势种为褐菖鲉Sebastiscus marmoratus和长鳍篮子鱼Siganus canaliculatus, 主要种有黄鳍棘鲷Acanthopagrus latus、条马鲾Equulites rivulatus、李氏䲗Callionymus richardsoni和真赤鲷Pagrus major等10种; 夏、秋季的最大优势种均为长鳍篮子鱼, 春、冬季的最大优势种分别为褐菖鲉和条马鲾。全湾鱼类年均资源密度为45836尾·km -2和 435.21kg·km -2, 资源密度具有明显的空间差异和季节变化。全年各站位种数变化范围在2~21种, 多样性指数H′在0.468~4.031, 丰富度指数d在0.120~1.926, 均匀度指数J在0.181~0.969。聚类分析、单因子相似性分析(ANOSIM)及相似性百分比(SIMPER) 分析表明该海域鱼类群落存在一定程度的时空异质性。相关性分析表明影响防城河口湾鱼类群落结构的主要环境因子为水温、盐度和硝酸盐。
何思璇 , 何斌源 . 防城河口湾鱼类群落结构及其与环境因子关系研究[J]. 热带海洋学报, 2019 , 38(5) : 86 -97 . DOI: 10.11978/2019001
Based on the survey in May, August and November 2016 and February 2017, the spatial and seasonal dynamics of the fish community structure and its relationship with the environmental factors in Fangchenghe Estuary of Guangxi, China were studied. A total of 95 species of fishes were collected in four seasons, belonging to two classes, 12 orders, 37 families, and 69 genera. Among them, 72 species were warm water fishes and 23 species were warm temperate fishes, accounting for 75.8% and 24.2% of the total, respectively. As for their ecological types, the bottom and near-bottom fishes were the dominant species, which accounted for 86.3% of the total species. In all, 60 species in spring, 59 species in summer, 54 species in autumn, and 46 species in winter were recorded. The similarity index between summer and autumn was the highest among season pairs. According to the index of relative importance (IRI), the whole-year dominant species were Sebastiscus marmoratus and Siganus canaliculatus, whose IRI values were higher than 500; 10 main species included Acanthopagrus latus, Equulites rivulatus, Callionymus richardsoni, Pagrus major, and so on, whose IRI values ranged from 100 to 500. Siganus canaliculatus acted as the biggest dominant species in both summer and autumn, so did Sebastiscus marmoratus in spring and Equulites rivulatus in winter. The annual average resource density and biomass of the study area were 45836 individual·km -2and 435.21 kg·km -2, respectively. The resource abundance varied significantly among pairs of seasons or regions. The scale of species abundance per sample ranged from 2 to 21 species; the species diversity index H′, from 0.468 to 4.031; the richness index d, from 0.120 to 1.926; and the evenness index J, from 0.181 to 0.969. Cluster analysis, analysis of similarity (ANOSIM) and similarity of percentage (SIMPER) showed that there existed some spatial-temporal heterogeneity in the fish community structure. Correlation analysis showed that the water temperature, salinity and nitrate content were the major environmental factors affecting the fish community structure.
表1 防城河口湾鱼类优势种及其相对重要值IRI (单位: %)Tab. 1 Dominant species and IRI values (units: %) of fish communities in Fangchenghe Estuary |
| 种 类 | 春 | 夏 | 秋 | 冬 | 周年 |
|---|---|---|---|---|---|
| 褐菖鲉Sebastiscus marmoratus | 6184.83 | 90.72 | 64.01 | 65.29 | 1592.22 |
| 长鳍篮子鱼Siganus canaliculatus | 66.70 | 1501.39 | 847.94 | 108.15 | 628.84 |
| 黄鳍棘鲷Acanthopagrus latus | 134.21 | 891.46 | 645.73 | 685.55 | 474.57 |
| 条马鲾Equulites rivulatus | 122.54 | 124.17 | 355.10 | 2140.66 | 375.19 |
| 真赤鲷 Pagrus major | 1625.70 | 161.38 | 3.46 | / | 323.73 |
| 李氏䲗 Callionymus richardsoni | 772.46 | 66.03 | 68.24 | 79.14 | 321.84 |
| 多鳞鱚Sillago sihama | 392.25 | 62.43 | 125.24 | 985.59 | 306.39 |
| 日本银鲈Gerres japonicus | 50.97 | 12.16 | 12.00 | 1352.86 | 173.41 |
| 鲬Platycephalus indicus | 74.28 | 138.30 | 317.88 | 144.40 | 138.94 |
| 二长棘犁齿鲷 Evynnis cardinalis | 539.79 | 7.90 | / | 23.51 | 119.65 |
| 斑头舌鳎Cynoglossus puncticeps | 76.97 | 448.16 | 19.39 | 80.75 | 117.18 |
| 高体斑鲆Pseudorhombus elevatus | 275.72 | 7.73 | 138.95 | 22.60 | 102.12 |
| 皮氏叫姑鱼Johnius belengerii | 37.59 | 172.12 | 573.08 | / | 92.67 |
| 粗高鳍鲉Vespicula trachinoides | 54.92 | 222.20 | 59.10 | 73.96 | 80.99 |
| 黑棘鲷Acanthopagrus schlegelii | 2.50 | 257.20 | 134.66 | 51.30 | 76.44 |
| 卵鳎Solea ovata | 1.94 | 137.25 | 426.65 | 78.47 | 71.42 |
| 中国花鲈Lateolabrax maculatus | 78.96 | 88.09 | 67.05 | 21.58 | 67.74 |
| 短吻鲾Leiognathus brevirostris | / | / | 8.72 | 505.31 | 34.13 |
| 鹿斑仰口鲾Secutor ruconius | / | 278.94 | 52.68 | 2.30 | 28.30 |
| 铅点多纪鲀Takifugu alboplumbeus | 15.44 | 37.45 | 30.74 | 38.14 | 26.64 |
| 条纹叫姑鱼Johnius fasciatus | / | 228.55 | 43.58 | / | 25.45 |
| 及达副叶鲹Alepes djedaba | 16.42 | 130.99 | 8.77 | / | 21.10 |
| 红尾银鲈Gerres erythrourus | 9.79 | / | 6.01 | 153.31 | 20.39 |
| 横带棘线鲬Grammoplites scaber | 21.95 | 63.98 | 2.68 | 1.80 | 16.72 |
| 斑纹舌鰕虎鱼Glossogobius olivaceus | 3.43 | / | 6.91 | 101.56 | 15.94 |
| 东方宽箬鳎 Brachirus orientalis | 32.67 | 21.60 | 1.50 | 9.34 | 15.94 |
| 日本钩嘴鳎 Heteromycteris japonicus | 21.88 | / | 2.10 | 68.47 | 12.67 |
| 长棘银鲈 Gerres filamentosus | 8.29 | / | 64.84 | 8.01 | 12.00 |
| 十刺银鲈Gerres decacanthus | / | / | 313.84 | / | 11.62 |
| 大头银姑鱼 Pennahia macrocephalus | 0.48 | 59.19 | 48.76 | / | 11.28 |
注:“/”表示未捕获。 |
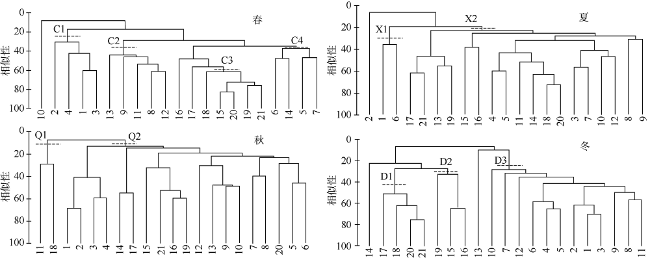
图5 4个季节防城河口湾鱼类群落等级聚类图C1~C4表示春季的4个显著差异的站位组, X1、X2表示夏季的2个显著差异的站位组, Q1、Q2表示秋季的2个显著差异的站位组, D1~D3表示冬季的3个显著差异的站位组。图中虚线表示某个组包括的站位范围 Fig. 5 The hierarchical cluster dendrogram of fish communities in Fangchenghe Estuary. C1~C4 represent the four significantly different station groups in spring. X1 and X2 represent the two significantly different station groups in summer. Q1 and Q2 represent the two significantly different station groups in autumn. D1~D3 represent the three significantly different station groups in winter. The dotted line in the figure represents the range of stations included in each group |
表2 防城河口湾各季节的鱼类群落数量指标与环境因子的相关性Tab. 2 Correlations between numerical indexes of fish community and their environmental factors in Fangchenghe Estuary |
| 季节 | 数量指标 | 显著正相关因子 | 显著负相关因子 |
|---|---|---|---|
| 春季 | J | 水温、溶解氧饱和度、无机磷 | 水深、盐度 |
| d | / | 悬浮物、COD、亚硝酸、盐硝酸盐、无机氮、总氮、硅酸盐 | |
| H′ | 溶解氧饱和度 | 悬浮物、COD、硝酸盐、氨氮、无机氮 | |
| S | / | COD、硝酸盐、无机氮、总氮 | |
| 生物量 | 盐度 | 水温、硝酸盐 | |
| 密度 | 盐度 | 水温、硝酸盐 | |
| 夏季 | J | 水温 | pH、溶解氧饱和度、BOD |
| d | 盐度 | / | |
| H′ | 水温、盐度 | COD | |
| S | 盐度 | 亚硝酸盐、硝酸盐、无机氮、总氮、无机磷、硅酸盐 | |
| 生物量 | pH、溶解氧饱和度、BOD | 水温 | |
| 密度 | pH、溶解氧饱和度、BOD | 水温 | |
| 秋季 | J | 水色、无机磷、总氮、总磷 | 透明度、硅酸盐 |
| d | 水深 | / | |
| H′ | / | / | |
| S | 透明度 | 水色、无机磷 | |
| 生物量 | / | / | |
| 密度 | / | / | |
| 冬季 | J | 盐度、悬浮物、pH、总磷 | 透明度 |
| d | / | BOD | |
| H′ | / | BOD、叶绿素a | |
| S | / | BOD | |
| 生物量 | / | 盐度、水温、pH | |
| 密度 | 亚硝酸盐、硝酸盐、无机氮、氨氮、总氮、无机磷、硅酸盐 | 水温、悬浮物、pH、盐度 |
注: “/”指无显著相关的环境因子 |
| [1] |
崔磊, 吕颂辉, 董悦镭 , 等, 2017. 围填海工程对淇澳岛附近水域环境因子与生物群落的影响[J]. 热带海洋学报, 36(2):96-105.
|
| [2] |
单秀娟, 陈云龙, 戴芳群 , 等, 2014. 黄海中南部不同断面鱼类群落结构及其多样性[J]. 生态学报, 34(2):377-389.
|
| [3] |
傅昕龙, 徐兆礼, 阙江龙 , 等, 2019. 北部湾西北部近海鱼类资源的时空分布特征研究[J]. 水产科学, 38(1):10-18.
|
| [4] |
管伟, 徐兆礼, 陈佳杰 , 2017. 福建南日岛南部水域鱼类群落结构及多样性[J]. 生态学报, 37(9):3172-3181.
|
| [5] |
广西海洋开发保护管理委员会, 1996. 广西海岛资源综合调查报告[M]. 南宁: 广西科学技术出版社.
Guangxi Oceanic Development and Protection Administration, 1996. Investigation report on the Guangxi sea island resources[M]. Nanning: Guangxi Science and Technology Press (in Chinese).
|
| [6] |
何斌源 , 1999. 广西两港湾红树林鱼类生态的比较研究[J]. 海洋通报, 18(1):28-35.
|
| [7] |
何斌源, 范航清, 莫竹承 , 2001. 广西英罗港红树林区鱼类多样性研究[J]. 热带海洋学报, 20(4):74-79.
|
| [8] |
何秀玲, 叶宁, 宣立强 , 2003. 雷州半岛红树林海区的鱼类种类调查[J]. 湛江海洋大学学报, 23(3):3-10.
|
| [9] |
胡聪 , 2014. 围填海开发活动对海洋资源影响评价方法研究[D]. 青岛: 中国海洋大学.
|
| [10] |
黄德练, 吴志强, 黄亮亮 , 等, 2013. 钦州港红树林鱼类群落时间变化格局及其与潮差等环境因子关系[J]. 桂林理工大学学报, 33(3):454-460.
|
| [11] |
赖廷和, 何斌源 , 2016. 广西北部湾海洋硬骨鱼类图鉴[M]. 北京: 科学出版社.
|
| [12] |
黎清华, 万世明, 何军 , 等, 2014. 近两百年来人类活动对北部湾潮间带环境的影响[J]. 海洋地质与第四纪地质, 34(1):57-64.
|
| [13] |
李显森, 梁志辉, 蒋明星 , 1987. 北部湾北部我国沿岸海区鱼类区系的初步调查[J]. 广西科学院学报, 3(2):95-116.
|
| [14] |
李显森, 于振海, 孙珊 , 等, 2013. 长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠[J]. 应用生态学报, 24(8):2353-2359.
|
| [15] |
李渊, 张静, 张然 , 等, 2016. 南沙群岛西南部海域和北部湾口海域底层游泳动物多样性[J]. 中国水产科学, 23(1):177-187.
|
| [16] |
陆健健 , 2003. 河口生态学[M]. 北京: 海洋出版社.
|
| [17] |
罗春业, 李英, 朱瑜 , 等, 1999. 广西北部湾鱼类区系的再研究[J]. 广西师范大学学报(自然科学版), 17(2):85-89.
|
| [18] |
求锦津, 王咏雪, 李铁军 , 等, 2018. 舟山长白海域主要游泳动物生态位及其分化研究[J]. 生态学报, 38(18):6759-6767.
|
| [19] |
邱永松 , 1988. 南海北部大陆架鱼类群落的区域性变化[J]. 水产学报, 12(4):303-313.
|
| [20] |
孙典荣 , 2008. 北部湾渔业资源与渔业可持续发展研究[D]. 青岛: 中国海洋大学.
|
| [21] |
孙鹏飞, 单秀娟, 吴强 , 等, 2014. 莱州湾及黄河口水域鱼类群落结构的季节变化[J]. 生态学报, 34(2):367-376.
|
| [22] |
王倩, 杨光, 吴孝兵 , 等, 2006. 广西合浦儒艮国家级自然保护区及邻近水域鱼类种数及保护对策[J]. 应用生态学报, 17(9):1715-1720.
|
| [23] |
王小林, 徐宾铎, 纪毓鹏 , 等, 2013. 海州湾及邻近海域冬季鱼类群落结构及其与环境因子的关系[J]. 应用生态学报, 24(6):1707-1714.
|
| [24] |
王雪辉, 邱永松, 杜飞雁 , 等, 2010. 北部湾鱼类群落格局及其与环境因子的关系[J]. 水产学报, 34(10):1579-1586.
|
| [25] |
王雪辉, 邱永松, 杜飞雁 , 等, 2011. 北部湾鱼类多样性及优势种的时空变化[J]. 中国水产科学, 18(2):427-436.
|
| [26] |
王雪辉, 林昭进, 杜飞雁 , 等, 2013. 南海西北部陆架区鱼类的种类组成与群落格局[J]. 生态学报, 33(7):2225-2235.
|
| [27] |
徐兆礼 , 2008. 瓯江口海域夏秋季鱼类多样性[J]. 生态学报, 28(12):5948-5956.
|
| [28] |
叶孙忠, 罗冬莲, 蔡建堤 , 等, 2018. 东山湾渔业生物群落结构及生物量分布特征[J]. 渔业研究, 40(5):358-365.
|
| [29] |
翟璐, 徐宾铎, 纪毓鹏 , 等, 2015. 黄河口及其邻近水域夏季鱼类群落空间格局及其与环境因子的关系[J]. 应用生态学报, 26(9):2852-2858.
|
| [30] |
张静, 陈永俊, 宋普庆 , 等, 2013. 福建东山湾游泳动物群落物种组成及其多样性[J]. 海洋渔业, 35(1):15-23.
|
| [31] |
张文超, 叶振江, 田永军 , 等, 2017. 北部湾洋浦海域鱼类群落结构[J]. 生态学杂志, 36(7):1894-1904.
|
| [32] |
中国海湾志编纂委员会, 1993. 中国海湾志第十二分册(广西海湾)[M]. 北京: 海洋出版社.
China Gulf Chronicle Compilation Committee, 1993. Editorial board of China bay survey (Vol. 12) Guangxi Bay[M]. Beijing: China Ocean Press (in Chinese).
|
| [33] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB 17378. 4-2007 海洋监测规范第4部分: 海水分析[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of China, 2008. GB 17378.4-2007 The specification for marine monitoring— Part 4: seawater analysis[S]. Beijing: Standards Press of China (in Chinese).
|
| [34] |
中华人民共和国农业部, 2013. SC/T 9403-2012 海洋渔业资源调查规范[S]. 北京: 中国农业出版社.
Ministry of Agriculture of the PRC, 2013. SC/T 9403-2012 Technical specification for marine fishery resources survey[S]. Beijing: China Agriculture Press (in Chinese).
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}