
安达曼海浮游有孔虫群落对全新世海洋环境变化的响应
|
张玲芝(1993—), 女, 江苏省连云港市人, 硕士研究生。E-mail: |
Copy editor: 林强
收稿日期: 2019-01-15
要求修回日期: 2019-04-01
网络出版日期: 2019-11-26
基金资助
国家自然科学基金项目(91228207)
国家自然科学基金项目(41476040)
中国科学院战略专项项目(XDA11030104)
版权
Response of planktonic foraminifera to Holocene marine environmental changes in the Andaman Sea
Copy editor: LIN Qiang
Received date: 2019-01-15
Request revised date: 2019-04-01
Online published: 2019-11-26
Supported by
National Natural Science Foundation of China(91228207)
National Natural Science Foundation of China(41476040)
Strategic Leading Science & Technology Programme, Chinese Academy of Sciences(XDA11030104)
Copyright
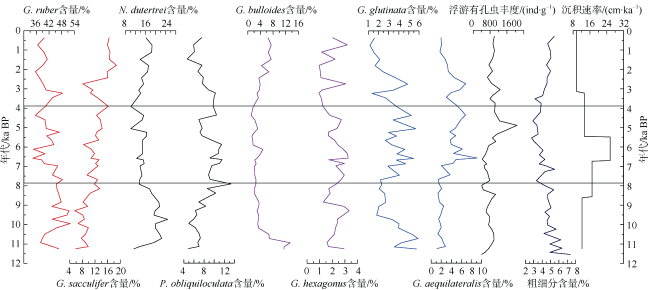
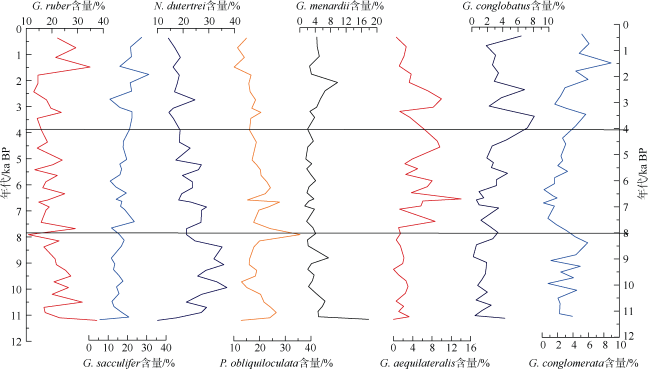
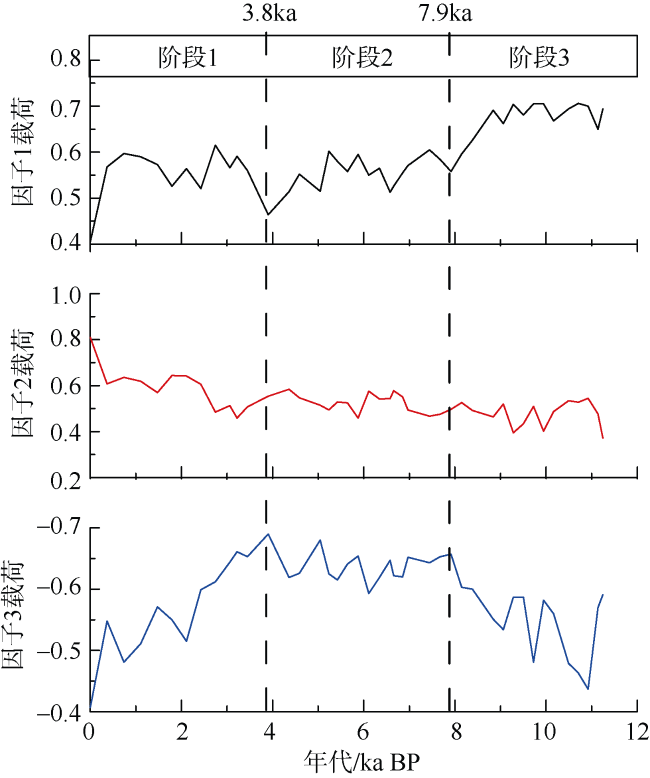
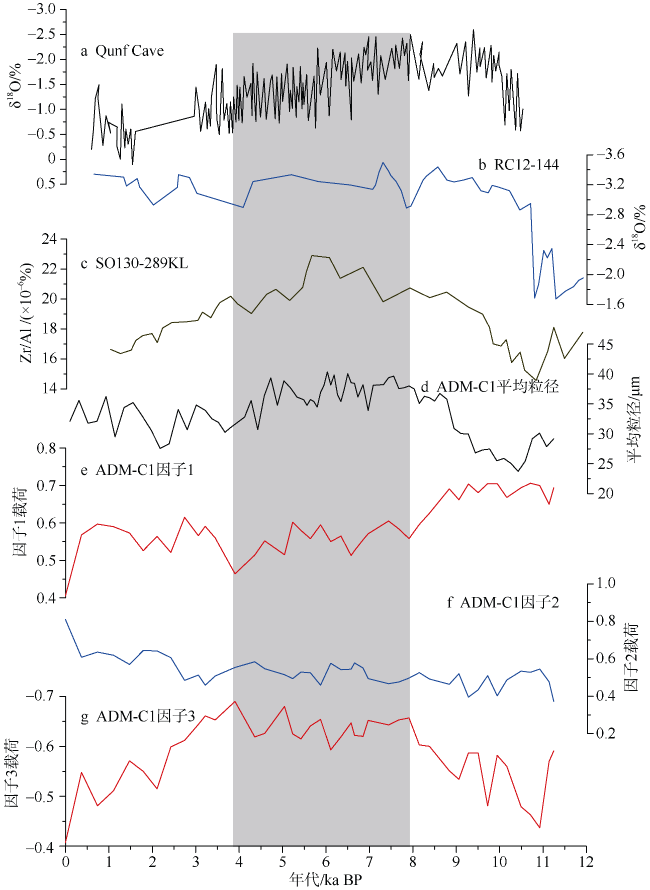
通过对安达曼海重力柱ADM-C1的浮游有孔虫群落分析, 探讨了该区全新世以来的海洋环境演化。研究发现, 该区浮游有孔虫群落总体以热带暖水种Globigerinoides ruber、Globigerinoides sacculifer、Neogloboquadrina dutertrei和Pulleniatina obliquiloculata等为主。其中G. ruber的相对丰度从早全新世至今呈逐渐降低的趋势, G. sacculifer则呈大致相反的变化趋势。N. dutertrei的相对丰度从11—7.9ka BP较高, 到7.9—3.8ka BP偏低, 3.8ka BP以来又逐渐升高。而P. obliquiloculata的变化趋势则与N. dutertrei大致相反。研究认为, 安达曼海不存在与冲绳海槽类似的晚全新世普林虫低值事件。浮游有孔虫群落Q型因子显示全新世海洋环境呈现三个明显的阶段变化: 早全新世 11—7.9ka BP期间, 浮游有孔虫群落以G. ruber、N. dutertrei与G. bulloides为主, 反映了该阶段较强的夏季风降水会导致表层水体盐度较低, 同时冬季风的影响相对较强; 7.9—3.8ka BP期间, 以G. ruber、P. obliquiloculata和Globigerinella aequilateralis为主, 指示水体盐度仍然较低, 同时次表层属种显著增多, 对应了该阶段强盛的印度夏季风, 上层海水混合强烈; 3.8—0ka BP期间, 以G. sacculifer、N. dutertrei和P. obliquiloculata为主要特征属种, 而G. ruber相对丰度明显下降, 表明该时期表层海水的盐度有显著上升, 对应了印度夏季风降水的明显减弱。安达曼海浮游有孔虫群落所呈现的全新世海洋环境阶段性变化和陆地夏季风记录有很好的一致性, 也与该孔敏感粒级组分所反映的印度季风的强度变化一致, 表明热带边缘海区的有孔虫群落组合可以很好地响应区域海洋环境变化。
张玲芝 , 向荣 , 唐灵刚 , 杨艺萍 , 钟福昌 . 安达曼海浮游有孔虫群落对全新世海洋环境变化的响应[J]. 热带海洋学报, 2019 , 38(6) : 51 -61 . DOI: 10.11978/2019010
Through analysis of planktonic foraminifera in a gravity core ADM-C1 from the Andaman Sea, the response of planktonic foraminifera to Holocene marine environmental changes in the sea was discussed. The result shows that the planktonic foraminiferal communities in this region are dominated by tropical warm-water species Globigerinoides ruber, Globigerinoides sacculifer, Neogloboquadrina dutertrei, and Pulleniatina obliquiloculata. The relative abundance of G. ruber shows a gradual decreased trend from the early Holocene to present, which is roughly opposite to that of G. sacculifer. The relative abundance of N. dutertrei is higher during 11 to 7.9 ka BP (before present), generally lower during 7.9 to 3.8 ka BP, and then gradually increases after 3.8 ka BP. P. obliquiloculata shows an almost opposite trend to that of N. dutertrei. Our result indicates no obvious Pulleniatina Minimum Eventduring the late Holocene, occurs in the Andaman Sea. Q-mode factor analyses of planktonic foraminifera in the core ADM-C1 identify three stages in marine environmental changes during the Holocene. During the early Holocene (11-7.9 ka BP), planktonic foraminifera assemblage was dominated by G. ruber, N. dutertrei and G. bulloides, which reflected that strong summer monsoon and precipitation led to lower sea surface salinity, while the influence of winter monsoon was also strong during this stage. During 7.9-3.8 ka BP, G. ruber, P. obliquiloculata and Globigerinella aequilateralis dominated, indicating lower sea surface salinity during this period. Meanwhile, there was obviously increase in abundance of subsurface water species, reflecting strong mixing in the upper-ocean waters, possibly due to the strong Indian Ocean summer monsoon at this stage. After 3.8 ka BP, G. sacculifer, N. dutertrei and P. obliquiloculata became the dominant species. The abundance of G. ruber declined obviously, indicating that the salinity of surface sea water significantly increased during this period, which was consistent with the reduced Indian Ocean summer monsoon precipitation. The phase change of the Holocene marine environment reflected by the planktonic foraminiferal assemblages in the Andaman Sea is well consistent with the terrestrial records on the Indian Ocean summer monsoon, and it also shows good consistent with the wind intensity variation in Indian Ocean summer monsoon reflected by sensitive grain size proxy of the same core, suggesting rapid response of planktonic foraminifera to marine environmental changes in the tropical marginal seas.
图1 研究区海流模式与取样站位(改自Cao et al, 2015)Fig. 1 Circulation patterns of the study area and sampling sites. Modified after Cao et al (2015) |
表1 ADM-C1孔中浮游有孔虫方差最大因子得分Tab. 1 Varimax factor score matrix for the planktonic foraminifera in Core ADM-C1(The bold numbers are species with higher scores) |
| 属种 | 主因子1 | 主因子2 | 主因子3 |
|---|---|---|---|
| Globigerinoides ruber | 2.943 | 0.466 | -3.503 |
| Globigerinoides sacculifer | -1.779 | 3.571 | -0.606 |
| Neogloboquadrina dutertrei | 2.458 | 2.370 | 1.926 |
| Pulleniatina obliquiloculata | -1.020 | 1.375 | -1.210 |
| Globorataloides hexagonus | 0.176 | -0.033 | -0.202 |
| Globigerinita glutinata | 0.154 | -0.015 | -0.411 |
| Globigerina bulloides | 1.371 | 0.882 | 1.496 |
| Globigerina calida | -0.110 | -0.034 | -0.389 |
| Globiberinella aequilateralis | -0.953 | 0.443 | -1.155 |
| Globigerinoides tenellus | -0.145 | 0.020 | -0.223 |
| Globoquadrina conglomerata | -0.128 | 0.415 | 0.119 |
| Globigerinoides conglobatus | -0.315 | 0.301 | -0.201 |
| Globorotalia menardii | -0.024 | 0.488 | 0.248 |
| Globorotalia scitula | 0.075 | -0.021 | 0.040 |
| Orbulina universa | -0.289 | 0.172 | -0.271 |
| Sphaeroidinella dehiscens | -0.072 | 0.131 | 0.009 |
| Globorotalia tumida | 0 | 0.035 | 0.028 |
| Globorotalia ungulata | 0.028 | 0.020 | -0.001 |
| Globorotalia theyeris | -0.004 | -0.008 | -0.042 |
| Candeina nitida | -0.036 | 0.025 | -0.039 |
| Globigerina digitata | -0.011 | 0.012 | -0.003 |
| Globigerinella adamis | -0.011 | 0.012 | -0.003 |
| 方差 | 36.253 | 28.209 | 34.714 |
| 累计方差 | 36.253 | 64.462 | 99.176 |
注: 加粗数字为得分较高属种 |
图5 ADM-C1孔中浮游有孔虫群落全新世阶段变化与其他古环境记录对比a. Qunf cave石笋δ18O值变化(Fleitmann et al, 2003); b. 安达曼海RC12-144孔G. ruber δ18O值变化(Rashid et al, 2007); c. SO130-289KL的Zr/Al(Deplazes et al, 2014); d. 主要粒组1.5~11.9μm与11.9~74μm的平均粒径 (黄云 等, 2017) ; e—g. ADM-C1孔中浮游有孔虫因子载荷变化。阴影部分为阶段2 Fig. 5 Changes of planktonic foraminiferal in Core ADM-C1 compared with other paleoenvironmental records. a. The records of speleothem δ18O from the Qunf Cave (Fleitmann et al, 2003). b. δ18Osw of Core RC12-344 in the Andaman Sea (Rashid et al., 2007); c. Zr/Al of SO130-289 KL (Deplazes et al, 2014); d. the average grain size variation of grain groups 1.5~11.9 m and 11.9~74 m in ADM-C1 (Huang et al, 2017); e-g. the variation of main planktonic foraminiferal factors in Core ADM-C1. The dark shade indicates phrase 2 |
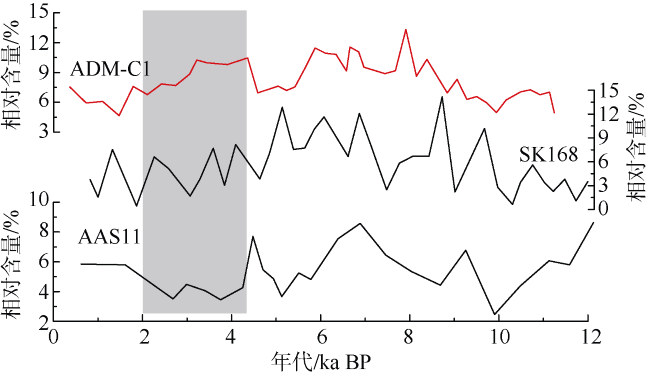
图6 安达曼海三个岩芯ADM-C1、SK168和AAS11 (Sijinkumar et al, 2011)的P. obliquiloculata相对含量对比阴影为普林虫低值事件Fig. 6 Abundance of P.obliquiloculata in Core ADM-C1, SK168 and AAS11 (Sijinkumar et al, 2011) |
| 1 |
曹鹏 , 2015. 末次盛冰期以来安达曼海东南部沉积特征及其对印度季风的响应[D]. 青岛: 中国海洋大学.
|
| 2 |
陈荣华, 郑玉龙 ,
|
| 3 |
丁旋, 方念乔 , 2006. 东北印度洋区BAR9427岩心末次冰期以来的古季风活动记录[J]. 地球科学—中国地质大学学报, 31(6):765-772.
|
| 4 |
方念乔, 陈学方, 丁旋 , 等, 2001. 孟加拉湾和东径90°海岭260ka以来的古海洋学记录与印度季风的影响[J]. 中国科学 D辑: 地球科学, 31(S1):280-286.
|
| 5 |
黄云, 向荣, 刘升发 , 等, 2017. 安达曼海沉积物粒度记录的全新世印度洋夏季风演化[J]. 热带海洋学报, 36(6):19-26.
|
| 6 |
黄云 , 2017. 安达曼海全新世以来高分辨率印度洋夏季风演化[D]. 北京: 中国科学院大学.
|
| 7 |
李保华, 王晓燕 , 2009. 末次冰期以来中国海区浮游有孔虫Pulleniatina obliquiloculata含量变化的地层学意义[J]. 微体古生物学报, 26(4):313-322.
|
| 8 |
向荣, 陈木宏, 张兰兰 , 等, 2010. 南海北部秋季活体浮游有孔虫的组成与分布[J]. 地球科学—中国地质大学学报, 35(1):1-10.
|
| 9 |
向荣, 刘芳, 孙有斌 , 等, 2011. 冲绳海槽末次冰消期以来的浮游有孔虫群落变化及5种古温度转换函数对比[J]. 地球科学—中国地质大学学报, 36(4):599-609.
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}