
干出状态下坛紫菜叶状体表面水膜与失水对光合作用光化学特性的影响
|
蔡逸洵(1996—), 女, 广东省汕头市人, 硕士研究生, 研究方向为海洋环境生态学。email: |
Copy editor: 姚衍桃
收稿日期: 2020-01-16
要求修回日期: 2020-02-29
网络出版日期: 2020-02-29
基金资助
广东省科技计划项目(2018B030311029)
广东省科技计划项目(2019B121202001)
版权
Effects of surface water film and desiccation on chlorophyll fluorescence characteristics of emersed Pyropia haitanensis thalli
Received date: 2020-01-16
Request revised date: 2020-02-29
Online published: 2020-02-29
Supported by
Science and Technology Program of Guangdong Province(2018B030311029)
Science and Technology Program of Guangdong Province(2019B121202001)
Copyright
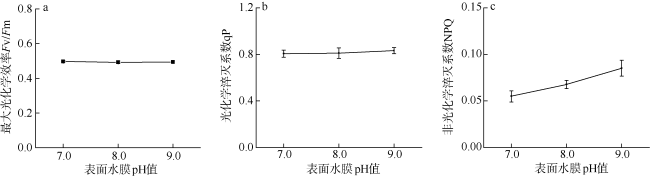
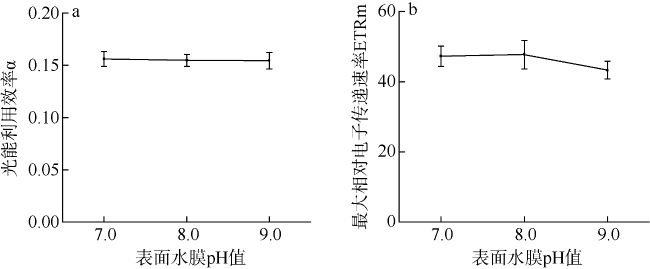
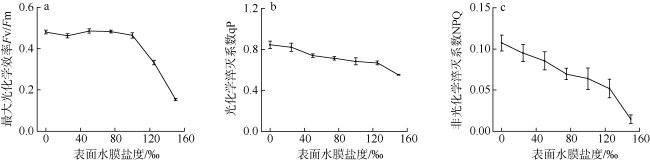
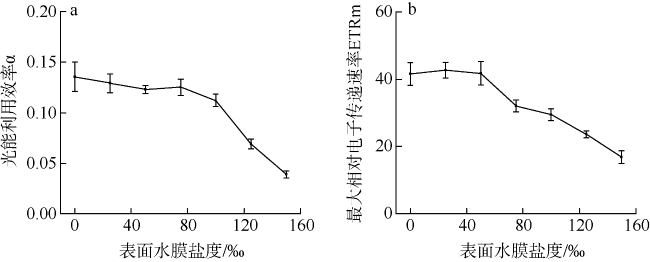
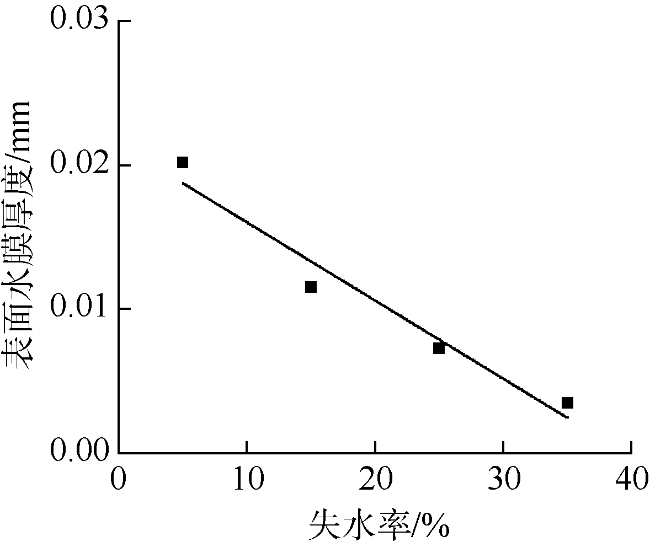
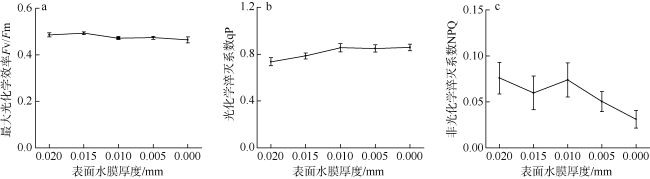
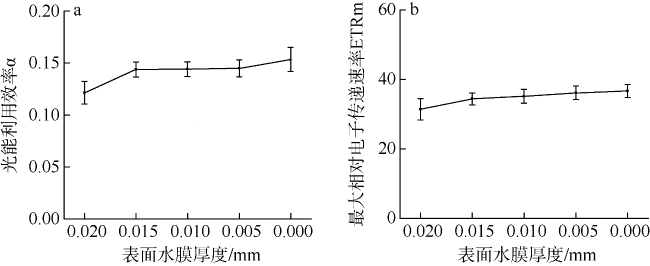
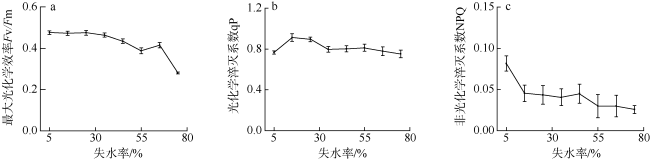
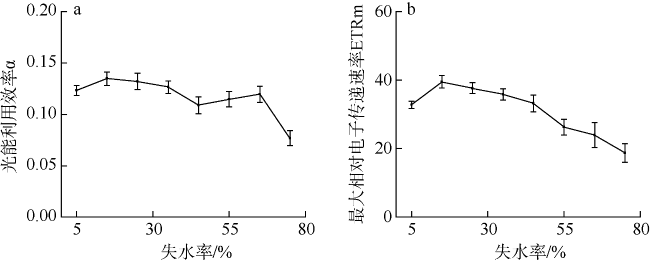
采用筏式吊养的大型经济海藻坛紫菜(Pyropia haitanensis)在栽培生长期间, 常周期性地处于干出状态。本文通过测定不同水膜状态下坛紫菜的叶绿素荧光动力学相关系数, 探讨了干出状态下坛紫菜叶状体表面水膜的理化特性与失水对其光合作用光化学特性的影响。实验数据显示, 干出初始状态下, 叶状体表面水膜pH值的升高对坛紫菜的最大光化学效率Fv/Fm、光化学淬灭系数qP、非光化学淬灭系数NPQ、最大相对电子传递速率ETRm和光能利用效率α均无显著性影响。当坛紫菜叶状体表面水膜盐度为0~75‰时, Fv/Fm、qP、NPQ、ETRm和α均保持稳定; 当叶状体表面水膜盐度增加到75‰~150‰时, Fv/Fm、NPQ、α和ETRm则显著下降, 较盐度为33‰时分别下降了66%、84%、70%和60%, 表明覆盖高盐度水膜的叶状体PSⅡ反应中心活性受到抑制。坛紫菜叶状体在失水初期的光合活性不受影响, 当叶状体表面水膜厚度降到0后, 叶状体细胞内部开始失水, 光合活性也显著下降。严重脱水状态(失水率为75%)下, Fv/Fm、NPQ、α和ETRm较失水初期分别下降了41%、69%、38%和43%。这些研究结果表明, 坛紫菜叶状体在干出失水初期, 水分损失是由于表面水膜损失引起的, 水膜厚度随着失水率的增加呈线性下降, 表面水膜厚度的下降影响了藻体的热耗散能力; 随着失水率进一步增加, 叶状体的光合活性和热耗散能力受到不利影响, 其中热耗散能力对叶状体含水量的变化更为敏感。
蔡逸洵 , 温嘉怡 , 邹定辉 . 干出状态下坛紫菜叶状体表面水膜与失水对光合作用光化学特性的影响[J]. 热带海洋学报, 2021 , 40(1) : 82 -90 . DOI: 10.11978/2020010
Pyropia haitanensis is an economically important seaweed species, usually cultured by means of raft suspension. The thallus are periodically exposed to air and face desiccation when the tide is low. In this study, we investigated the effects of physicochemical properties of surface water film and desiccation on chlorophyll fluorescence characteristics in emersed thallus of Pyropia haitanensis. The results showed that the increase of the pH value of the thallus surface water film had no significant effect on the maximum quantum yield (Fv/Fm) of Photosystem Ⅱ(PSⅡ), photochemical quenching (qP) and non-photochemical quenching (NPQ), as well as the light utilize efficiency (α) and relative maximum photosynthetic electron transport rate (ETRm). The Fv/Fm, qP, NPQ, ETRm, and α remained stable over the salinity range of 0~75‰. However, when the salinity reached 75‰~150‰, Fv/Fm, NPQ, ETRm, and α declined significantly. High salinity in the thallus surface water film reduced Fv/Fm, NPQ, α, and ETRm by 66%, 84%, 70%, and 60%, respectively. This suggests that the activity of PSII reaction center was inhibited by high salinity in the thallus surface water film. The thallus photosynthetic activity remained stable during the initial stage of water loss. When the thallus surface water film thickness dropped to 0, water was lost within the thallus cells and the photosynthetic activity decreased progressively. Severe dehydration of 75% reduced the Fv/Fm, NPQ, α and ETRm by 41%, 69%, 38% and 43%, respectively. The results showed that the thallus surface water film thickness decreased linearly with the increase of initial water loss. With further water loss, the thallus photosynthetic activity and heat dissipation capacity were negatively affected, and the heat dissipation capacity was more sensitive to the change of water content of algal bodies.
图6 不同水膜厚度下坛紫菜最大光化学效率Fv/Fm(a)、光化学淬灭系数qP(b)和非光化学淬灭系数NPQ(c)的变化Fig. 6 Changes in the maximum quantum yield of Photosystem Ⅱ(a), photochemical quenching (b) and non-photochemical quenching (c) in different thicknesses of thallus surface water film under dehydration |
| [1] |
姜真杰, 2017. 基于PS的植物叶片表面积的测算[J]. 农业与技术, 37(14):41-42 (in Chinese).
|
| [2] |
钱飞箭, 2014. 坛紫菜受高温和干出胁迫的机理及脂代谢关键基因的研究[D]. 宁波: 宁波大学.
|
| [3] |
滕飞, 陈玉婷, 徐燕, 等, 2016. 用于坛紫菜光合效率的DIVING-PAM参数优化研究[J]. 集美大学学报(自然科学版), 21(2):81-86.
|
| [4] |
张忠山, 王晓梅, 刘峰, 等, 2018. 紫菜抗逆境生理与调控机制研究进展[J]. 植物生理学报, 54(6):960-972.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}