
Khai岛和Pathiu岛珊瑚礁沉积物细菌多样性及细菌粗提物延缓秀丽隐杆线虫衰老活性研究*
|
韩敏敏(1993—), 女, 山东省济宁市人, 硕士研究生, 主要从事海洋药物基础与产品开发研究。E-mail: |
Copy editor: 殷波
收稿日期: 2019-12-05
要求修回日期: 2020-03-03
网络出版日期: 2020-03-05
基金资助
国家自然科学基金项目(41566004)
国家自然科学基金项目(21662006)
广西中医药大学岐黄工程高层次人才团队培育项目(2018006)
广西中医药大学博士科研启动基金项目(2017BS039)
广西中医药大学海洋药物研究院团队科研专项经费项目(2018ZD005-A06)
版权
Studies on bacterial diversity in coral reef sediments in Khai Island and Pathiu Island and bacterial crude extract retards aging activity of Caenorhabditis elegans*
Copy editor: YIN Bo
Received date: 2019-12-05
Request revised date: 2020-03-03
Online published: 2020-03-05
Supported by
National Natural Science Foundation(41566004)
National Natural Science Foundation(21662006)
Development Program of High-level Talent Team under Qihuang Project of Guangxi University of Chinese Medicine(2018006)
Doctoral Research Foundation of Guangxi University of Traditional Chinese Medicine(2017BS039)
Scientific Research Foundation of Institute of Marine Drugs, GUCM(2018ZD005-A06)
Copyright
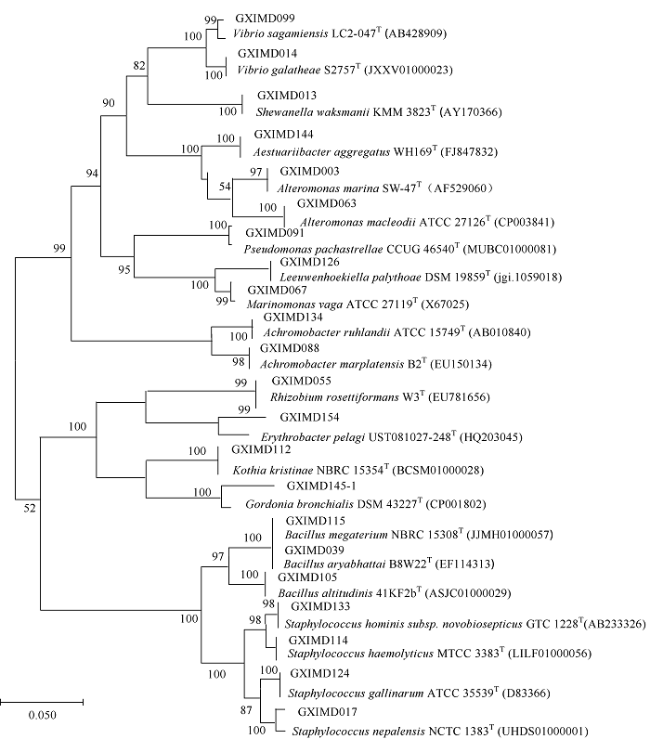
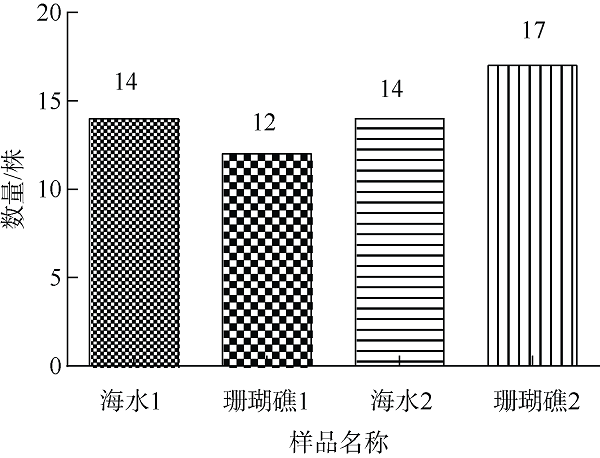
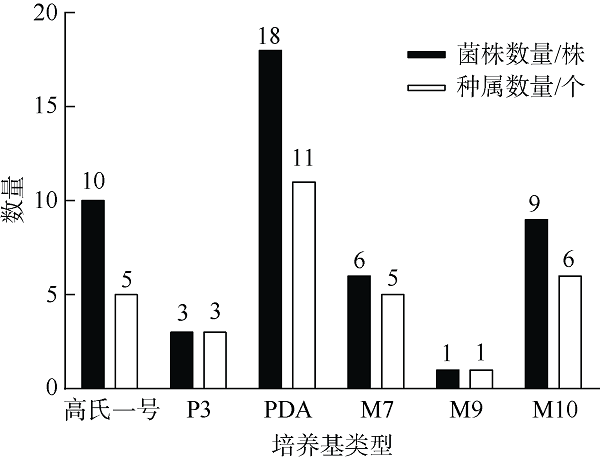
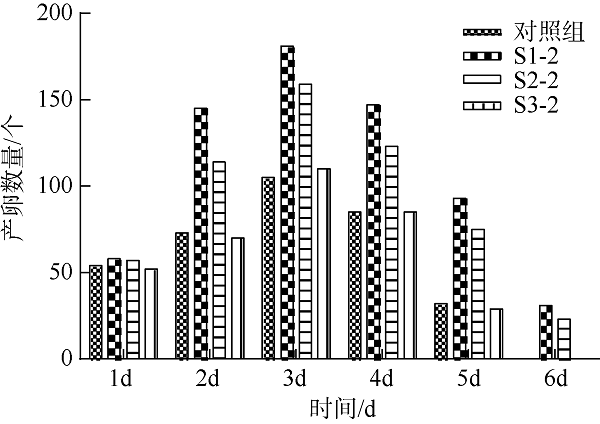
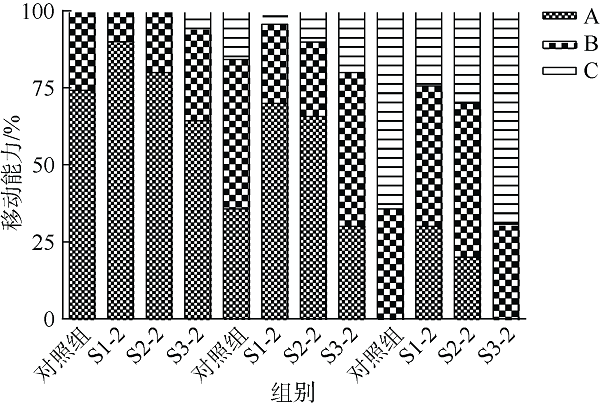
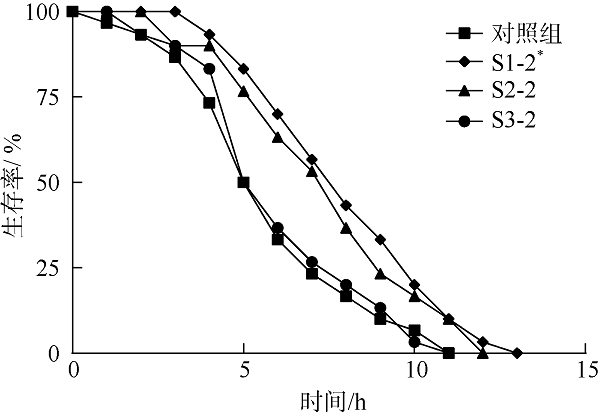
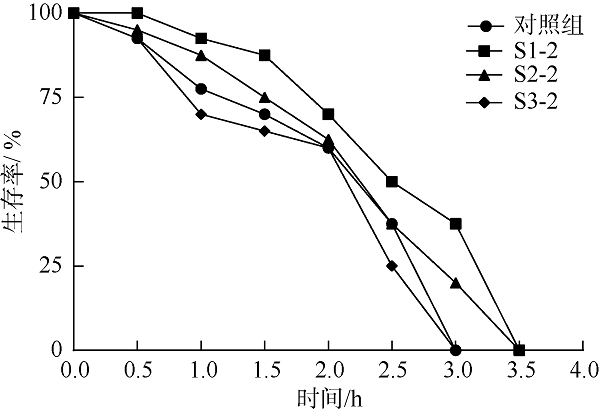
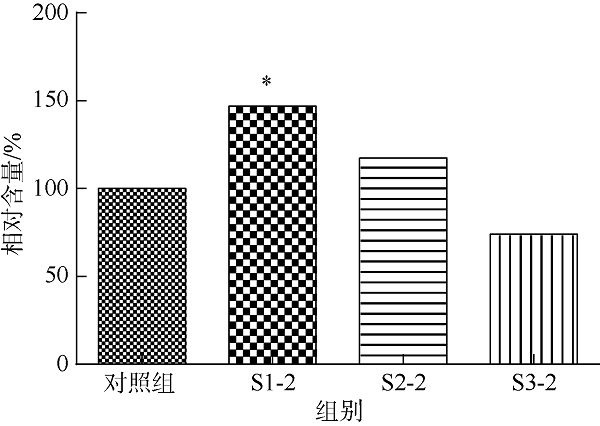
在Khai岛和Pathiu岛珊瑚礁生长海域采集2份珊瑚礁沉积物样品, 将样品预处理后采用6种分离培养基进行分离培养, 获得活性海洋细菌菌株, 通过PCR扩增和16s rRNA基因序列测定, 分析物种多样性, 并构建系统发育树。以模式生物秀丽隐杆线虫为模型考察菌株发酵提取物延缓衰老的作用。实验结果表明, 从4种样品中分离鉴定出22株细菌, 分别属于11目13科14属。16S rRNA基因序列相似性低于98.65%的细菌有2株, 疑似为潜在新种。GXIMD014 (Vibrio galatheae)和GXIMD112 (Kocuria kristinae)的500μg·mL-1细菌粗提物具有延缓线虫衰老的活性, 能不同程度的延长线虫寿命, 提高线虫产卵量、运动能力、热激损伤能力、抗氧化损伤能力、体内酶活性以及改善虫体表皮形态。Khai岛和Pathiu岛珊瑚礁沉积物细菌具有较高的物种多样性, 蕴藏着丰富的微生物资源, 且部分细菌菌株具有延缓衰老的生物活性。
韩敏敏 , 李蜜 , 刘昕明 , 刘永宏 , 龙超 , 钟振国 , 易湘茜 , 高程海 . Khai岛和Pathiu岛珊瑚礁沉积物细菌多样性及细菌粗提物延缓秀丽隐杆线虫衰老活性研究*[J]. 热带海洋学报, 2020 , 39(5) : 19 -29 . DOI: 10.11978/2019126
Two coral reef sediment samples were collected from the coral reef growth areas of Khai Island and Pathiu Island After the samples were pretreated, six kinds of separation medium were adopted to obtain the active Marine bacterial strains. Through PCR amplification and 16s rRNA gene sequencing, species diversity was analyzed and phylogenetic trees were constructed. The effect of fermentation extract of Caenorhabditis elegans strain on senescence was investigated. The results showed that 22 strains of bacteria were isolated and identified from four samples, belonging to 11 orders, 13 families and 14 genera. There were two strains of bacteria with 16S rRNA gene sequence similarity less than 98.65 %, which were suspected to be potential new species. The crude extracts of 500 μg·mL -1bacteria from GXIMD014 (Vibrio galatheae) and GXIMD112 (Kocuria kristinae) have the activity of delaying the aging of Caenorhabditis elegans, and can prolong the life of Caenorhabditis elegans to different degrees, improve the oviposition, motility, heat-induced injury, ability to resist oxidative damage, enzyme activity in vivo, and improve the epidermal morphology of Caenorhabditis elegans. Coral reef sediment bacteria of Khai Island and Pathiu Island have high species diversity and rich microbial and bacterial resources, and some bacterial strains have anti-aging biological activity.
Key words: coral reef; marine bacteria; species diversity; anti-aging activity
表1 22株珊瑚礁沉积物细菌的物种多样性Tab. 1 The species diversity of 22 sediment bacteria in coral reefs |
| 类群 | 菌株编号 | 相似性最高模式菌株 | 16S rRNA基因 序列相似性/% | GenBank登录号 |
|---|---|---|---|---|
| Kocuria sp. | GXIMD112 | K. Kristinae (BCSM01000028) | 99.74 | MT111730 |
| Vibrio sp. | GXIMD014 | V. Galatheae (JXXV01000023) | 99.35 | MT111742 |
| GXIMD099 | V. Sagamiensis (AB428909) | 98.75 | ||
| Bacillus sp. | GXIMD115 | B. Megaterium (JJMH01000057) | 99.74 | |
| GXIMD039 | B. Aryabhattai (EF114313) | 99.48 | ||
| GXIMD105 | B. Altitudinis (ASJC01000029) | 98.97 | ||
| Pseudomonas sp. | GXIMD091 | P. Pachastrellae (MUBC01000081) | 98.97 | |
| Shewanel sp. | GXIMD013 | S.Waksmanii (AY170366) | 99.59 | |
| Staphylococcus sp. | GXIMD124 | S. Gallinarum (D83366) | 99.23 | |
| GXIMD017 | S. Nepalensis (UHDS01000001) | 99.09 | ||
| GXIMD114 | S. Haemolyticus (LILF01000056) | 100.00 | ||
| GXIMD133 | S. hominis subsp. Novobiosepticus (AB233326) | 99.35 | ||
| Achromobacter sp. | GXIMD088 | A. marplatensis (EU150134) | 98.83 | |
| GXIMD134 | A. Ruhlandii (AB010840) | 99.61 | ||
| Leeuwenhoekiella sp. | GXIMD126 | L. Palythoae (jgi.1059018) | 98.83 | |
| Marinomonas sp. | GXIMD067 | M. Vaga (X67025) | 99.09 | |
| Gordonia sp. | GXIMD145-1 | G. Bronchialis (CP001802) | 99.23 | |
| Rhizobium sp. | GXIMD055 | R. Roseltifomans (EU781656) | 99.36 | |
| Erythrobacter sp. | GXIMD154 | E. Pelagi (HQ203045) | 99.87 | |
| Alteromonas sp. | GXIMD063 | A. Macleodii (CP003841) | 99.61 | |
| GXIMD003 | A. Marina (AF529060) | 97.93 | ||
| Aestuariibacter sp. | GXIMD144 | A. aggregatus (FJ847832) | 96.68 |
注: 2株活性菌株(GXIMD112和GXIMD014)的16S rRNA基因序列提交到GenBank数据库得到GenBank登录号, 其他菌株未获得GenBank登录号 |
表2 22株珊瑚礁沉积物细菌的粗提物对秀丽隐杆线虫生存寿命的影响($\bar{x}$±s, n=40)Tab. 2 Effects of crude extracts from 22 coral reef sediment bacteria on the lifespan of Caenorhabditis elegans ($\bar{x}$±s, n=40) |
| 菌株编号 | 平均寿命/d | 最长寿命/d | 菌株编号 | 平均寿命/d | 最长寿命/d |
|---|---|---|---|---|---|
| 1% DMSO | 16.68±1.057 | 18.83 | GXIMD039 | 19.75±1.060* | 21.89 |
| GXIMD055 | 14.50±0.844 | 16.21 | GXIMD114 | 15.30±0.834 | 16.99 |
| GXIMD099 | 15.50±0.938 | 17.40 | GXIMD115 | 14.25±0.839 | 15.95 |
| GXIMD154 | 16.56±1.126 | 18.84 | GXIMD091 | 17.10±1.261 | 19.65 |
| GXIMD144 | 14.30±0.818 | 15.96 | GXIMD088 | 15.65±0.826 | 17.32 |
| GXIMD112 | 20.75±1.417** | 23.62 | GXIMD145-1 | 14.27±0.891 | 16.08 |
| GXIMD013 | 16.20±0.891 | 18.00 | GXIMD014 | 21.75±1.119** | 22.01 |
| GXIMD124 | 16.45±1.041 | 18.56 | GXIMD003 | 15.05±0.950 | 16.97 |
| GXIMD017 | 17.22±1.250 | 19.78 | GXIMD134 | 16.10±1.107 | 18.34 |
| GXIMD126 | 15.20±0.925 | 17.07 | GXIMD063 | 13.25±0.867 | 15.00 |
| GXIMD067 | 13.15±0.671 | 14.51 | GXIMD133 | 14.40±0.817 | 16.05 |
| GXIMD105 | 14.22±1.010 | 16.26 |
注: *表示与对照组比较, 差异显著(p<0.05); **表示与对照组比较, 差异极显著(p<0.01) |
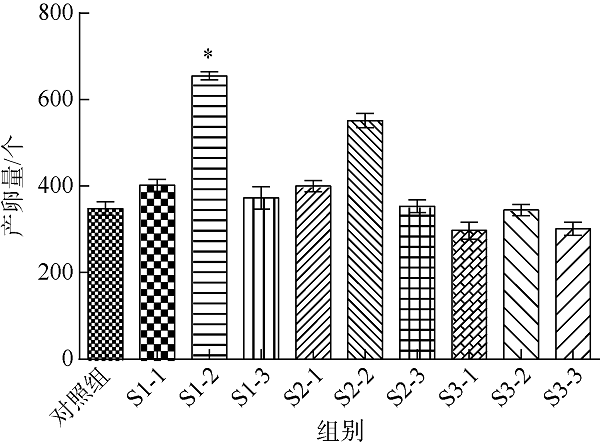
表3 3株细菌粗提物样品的基本信息Tab. 3 Basic information of crude extract samples of three strains of bacteria |
| 属 | 种 | 菌株粗提物 | 菌株粗提物编号 | 浓度/(μg·mL-1) | 菌株粗提物编号 |
|---|---|---|---|---|---|
| Vibrio sp. | Vibrio galatheae | GXIMD014细菌粗提物 | S1 | 250 | S1-1 |
| 500 | S1-2 | ||||
| 1000 | S1-3 | ||||
| Kocuria sp. | Kocuria kristinae | GXIMD112细菌粗提物 | S2 | 250 | S2-1 |
| 500 | S2-2 | ||||
| 1000 | S2-3 | ||||
| Bacillus sp. | Bacillus aryabhattai | GXIMD039细菌粗提物 | S3 | 250 | S3-1 |
| 500 | S3-2 | ||||
| 1000 | S3-3 |
图10 样品组与对照组线虫的扫描电镜图(3000×)a. 对照组, 图中箭头所指为成虫表皮出现的老化现象; b和c. S1和S2, 成虫未见表皮结构异常; d. S3, 图中箭头所指为成虫表皮有轻微“褶皱”现象 Fig. 10 Scanning electrogram of Caenorhabditis elegans in sample group and control group (3000×). (a) Control group, the arrows in the figure refer to the aging phenomenon of the adult. (b and c) S1 and S2, no epidermal structural abnormalities were observed in the adults. (d) S3, the arrow in the figure indicates that the epidermis of the adult has a slight “fold” |
| [1] |
蔡雪芹, 2018. 浅谈珊瑚及其生态功能与保护[J]. 海洋与渔业, (7):100-102 (in Chinese).
|
| [2] |
李菲, 高程海, 余炼, 等, 2018. 川蔓藻内生及根际细菌多样性与抑菌活性研究[J]. 广西植物, 38(7):922-933.
|
| [3] |
孙静, 王素英, 张德超, 等, 2014. 海南红树林根系土壤中可培养细菌的多样性分析[J]. 海洋科学, 38(7):27-33.
|
| [4] |
王怀玲, 2018. 蓝莓多酚化合物抗衰老活性及作用机制研究[D]. 广州: 华南理工大学: 1-172.
|
| [5] |
周双清, 黄小龙, 黄东益, 等, 2010. Chelex-100快速提取放线菌DNA作为PCR扩增模板[J]. 生物技术通报, 22(2):123-125.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}