
南海西北部夏冬季浮游介形类的分布及其影响因素
作者简介:廖彤晨(1995—), 女, 广东省佛山市人, 硕士, 研究方向是浮游动物生态学。E-mail: liaotongchen@foxmail.com
Editor: 殷波
收稿日期: 2019-05-20
要求修回日期: 2019-07-13
网络出版日期: 2020-03-10
基金资助
国家自然科学基金项目(41576125)
国家科技基础资源调查专项(2017FY201404)
广东省科技计划项目(2018B030320005)
版权
Distribution of planktonic ostracods and its influencing factors during summer and winter in the northwestern coastal waters of South China Sea
Received date: 2019-05-20
Request revised date: 2019-07-13
Online published: 2020-03-10
Supported by
National Natural Science Foundation of China(41576125)
Science and Technology Basic Resources Investigation of China(2017FY201404)
Science and Technology Planning Projects of Guangdong Province of China(2018B030320005)
Copyright
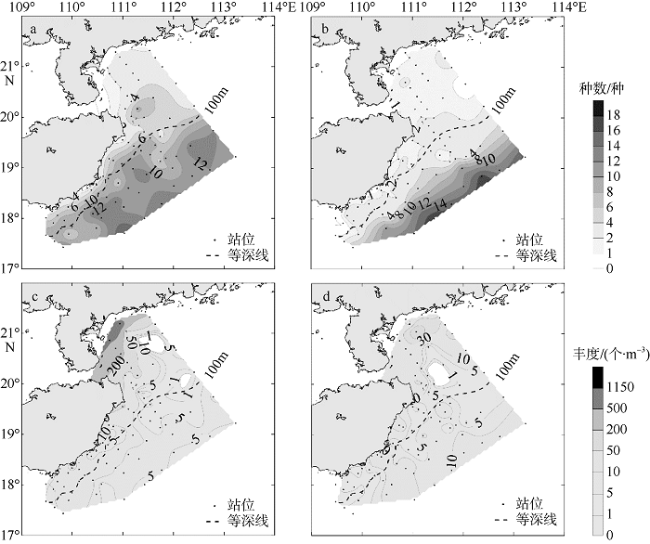
海洋浮游介形类(Ostracods)是一类分布较广的小型甲壳动物, 在海洋碳循环中起重要作用。目前国内海洋浮游动物采样常使用505μm网目孔径的浮游生物网, 从而导致个体较小的介形类被忽略。文章根据2006年7—8月(夏季)和2006年12月—2007年1月(冬季)在南海西北部海域分别使用网目孔径为505μm和160μm浮游生物网采集的样品, 比较不同网目所获浮游介形类种类和丰度的差异, 分析介形类种类和丰度的分布特征, 探讨环境因素对其时空分布的影响。160μm网目采集的浮游介形类种类和丰度均高于505μm。基于160μm网目数据分析浮游介形类群落结构: 1) 鉴定浮游介形类32种, 其种数由近岸向外海递增, 等深线100m以浅海域的种数显著高于100m以深海域(p<0.01); 2) 夏季浮游介形类的丰度高于冬季, 夏季丰度高值区主要出现在雷州半岛东部和琼东近岸, 显著高于外海(p<0.05), 冬季介形类丰度分布较为均匀; 3) 优势种针刺真浮萤(Euconchoecia aculeata)是近岸高丰度的主要贡献种; 4) 介形类种数与温度和水深呈显著正相关, 丰度与叶绿素a浓度呈正相关, 而与温度和盐度呈负相关。夏季琼东沿岸上升流和粤西沿岸流有助于浮游介形类在雷州半岛和海南岛东部近岸海域形成较高的丰度, 最高达1252个·m-3。同时, 建议今后研究浮游介形类采用网目孔径小的浮游生物网进行采集以便全面评估其群落结构特征。
廖彤晨 , 尹健强 , 李开枝 , 谭烨辉 . 南海西北部夏冬季浮游介形类的分布及其影响因素[J]. 热带海洋学报, 2020 , 39(2) : 77 -87 . DOI: 10.11978/2019050
Marine planktonic ostracods are small crustaceans with a wide distribution, playing an important role in ocean carbon cycle. The ostracods in smaller size are usually underestimated when marine zooplankton samples were collected with 505 μm mesh. Based on the zooplankton samples collected with 505 and 160 μm meshes in the northern South China Sea from July to August 2006 (summer) and from December 2006 to January 2007 (winter), the capture efficiency for the ostracods using two planktonic mesh sizes was compared, and the community structure and the influence of environmental factors on their distributions were analyzed. We found that the abundance of ostracods obtained by the 160 μm mesh net was six times that by the 505 μm net in summer. The community structure of planktonic ostracods collected by the 160 μm mesh was analyzed with the following findings. 1) The species richness of planktonic ostracods of a total of 32 species was identified to increase from nearshore to offshore, and the difference between the areas with water depth shallower and deeper than 100 m was extremely significant (p<0.01). 2) The abundance of planktonic ostracods was higher in summer, when the high abundance area appeared mainly in the coastal waters of the Leizhou Peninsula and northeast Hainan Island higher than that in offshore significantly (p<0.05), and lower in winter, when the distribution of ostracods abundance was homogeneous. 3) Euconchoecia aculeata, dominant species, contributed the most to high abundance in the nearshore. 4) The species richness of ostracods was positively correlated with temperature and water depth; and the abundance was negatively correlated with temperature and salinity, and positively correlated with chlorophyll α concentration. Qiongdong coastal upwelling and Yuexi Coastal Current boosted the abundance of planktonic ostracods up to 1252 ind·m -3 in the coastal waters of the Leizhou Peninsula and northeast Hainan Island. Results suggest that smaller mesh net should be used for the overall assessment of community characteristics of planktonic ostracods.
表1 夏冬季南海西北部160μm和505μm网目孔径采集的浮游介形类种数、平均丰度和优势种比较Tab. 1 Comparison of species richness, abundance and dominant species of pelagic ostracods by 160 and 505 μm mesh sizes in the northwestern South China Sea |
| 夏季 | 冬季 | |||
|---|---|---|---|---|
| 160μm | 505μm | 160μm | 505μm | |
| 种数 | 29 | 28 | 31 | 23 |
| 平均丰度/(个·m-3) | 59.97±181.82 | 7.14±10.12 | 9.89±12.80 | 1.97±2.46 |
| 针刺真浮萤 | 49.58±179.88 (0.59) | 2.31±7.72 (0.09) | 6.95±12.54 (0.58) | 0.77±1.90 (0.18) |
| 后圆真浮萤 | 3.48±6.70 (0.03) | 1.73±3.33 (0.14) | 1.28±3.08 (0.07) | 0.62±1.30 (0.17) |
| 同心假浮萤 | 2.67±5.57 (0.03) | 1.23±2.03 (0.14) | - | 0.09±0.27 (0.01) |
| 尖突海萤 | - | 0.89±4.09 (0.03) | - | - |
| 齿形海萤 | - | - | - | 0.15±0.60 (0.01) |
注: 括号内为优势种丰度的优势度;“-”表示种类在对应的季节和网型中不是优势种 |
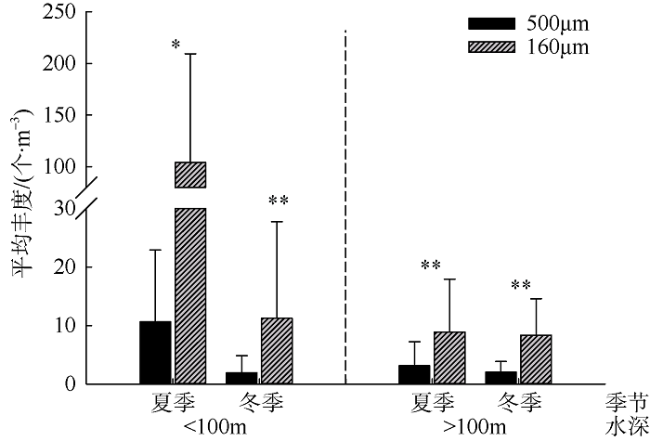
图3 南海西北部夏冬季160μm和505μm网目孔径采集浮游介形类丰度对比*表示p<0.05, 差异显著; **表示p<0.01, 差异极显著 Fig. 3 Comparison of sampling abundance between 160 and 505 μm meshes in the northwestern South China Sea. * represents p<0.05 and significant difference; ** represents p<0.01 and extremely significant difference |
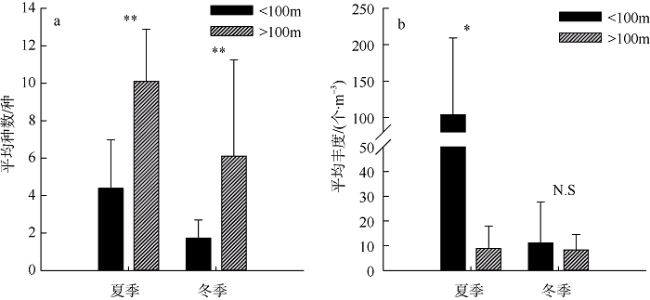
图4 浮游介形类平均种数(a)和平均丰度(b)变化*表示 p<0.05, 差异显著; **表示 p<0.01, 差异极显著; N.S表示差异不显著 Fig. 4 Variations of mean species richness (a) and abundance (b) of pelagic ostracods. * represents p<0.05 and significant difference; ** represents p<0.01 and extremely significant difference; N.S represents no significance |
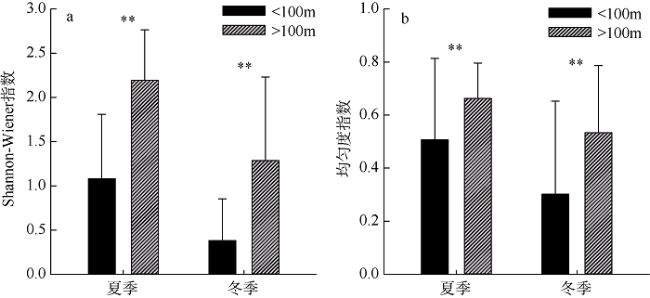
图5 夏冬季浮游介形类香农-威纳指数(a)和均匀度指数(b)的区域对比*表示 p<0.05, 差异显著; **表示 p<0.01, 差异极显著 Fig. 5 Comparison of Shannon-wiener index (a) and evenness index (b) of pelagic ostracods. * represents p<0.05 and significant difference; ** represents p<0.01 and extremely significant difference |
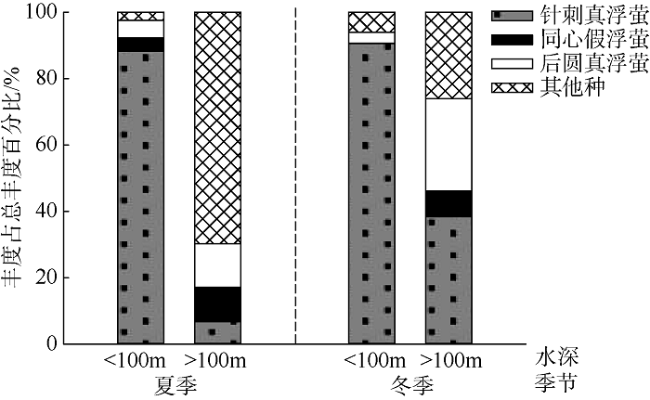
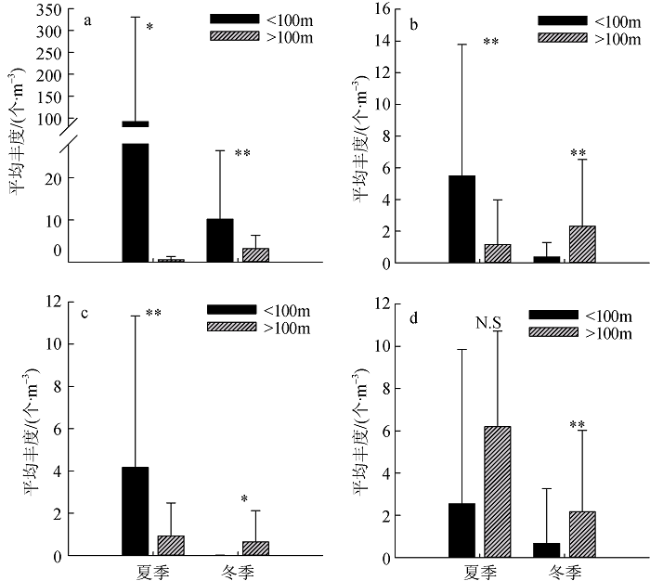
图8 优势种丰度的季节变化a. 针刺真浮萤; b. 后圆真浮萤; c. 同心假浮萤; d. 其他种类。*表示 p<0.05, 差异显著; **表示 p<0.01, 差异极显著; N.S表示差异不显著 Fig. 8 Variations of dominant and other species abundances in summer and winter. (a) Euconchoecia aculeata; (b) Euconchoecia maimai; (c) Pseudoconchoecia concentrica; and (d) other species. * represents p<0.05 and significant difference; ** represents p<0.01 and extremely significant difference; N.S represents no significance |
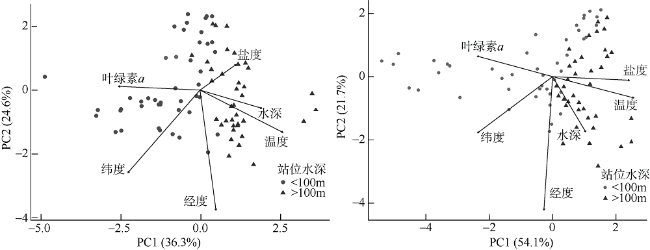
表2 南海西北部环境因子PCA主要计算结果Tab. 2 The main calculated results of environmental PCA in the northwestern continental shelf of the South China Sea |
| 夏季 | 冬季 | |||
|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | |
| 贡献率 | 0.36 | 0.25 | 0.54 | 0.22 |
| 累计贡献率 | 0.36 | 0.61 | 0.54 | 0.76 |
| 温度 | 0.53 | -0.27 | 0.51 | -0.13 |
| 盐度 | 0.23 | 0.16 | 0.49 | -0.02 |
| 叶绿素a浓度 | -0.53 | 0.02 | -0.48 | 0.13 |
| 经度 | 0.10 | -0.77 | -0.05 | -0.85 |
| 纬度 | -0.46 | -0.53 | -0.47 | -0.36 |
| 水深 | 0.40 | -0.12 | 0.21 | -0.35 |
图9 夏季(a)和冬季(b)浮游介形类群落结构PCA结果Fig. 9 PCA result of pelagic ostracods. (a) summer, and (b) winter |
表3 夏冬季浮游介形类和环境因子的Pearson相关系数rTab. 3 Pearson correlation coefficients r, between pelagic ostracods and environmental factors |
| 温度 | 盐度 | 水深 | 叶绿素a | ||
|---|---|---|---|---|---|
| 夏 季 | 丰度 | -0.33** | -0.37** | -0.12 | 0.39** |
| 种数 | 0.69** | 0.11 | 0.48** | -0.41** | |
| H' | 0.66** | 0.17 | 0.48** | -0.41** | |
| J | 0.37** | 0.23* | 0.27* | -0.25* | |
| 针刺真浮萤丰度 | -0.32** | -0.40** | -0.12 | 0.25* | |
| 后圆真浮萤丰度 | -0.26* | 0.05 | -0.15 | 0.18 | |
| 同心假浮萤丰度 | -0.34** | -0.31** | -0.16 | 0.45** | |
| 其他种类丰度 | 0.26* | -0.02 | 0.06 | -0.23* | |
| 冬 季 | 丰度 | -0.46** | -0.32** | -0.05 | 0.36** |
| 种数 | 0.47** | 0.16 | 0.70** | -0.31** | |
| H' | 0.51** | 0.23* | 0.60** | -0.33** | |
| J | 0.38** | 0.27* | 0.25* | -0.27* | |
| 针刺真浮萤丰度 | -0.60** | -0.44** | -0.15 | 0.50** | |
| 后圆真浮萤丰度 | 0.17 | 0.19 | -0.05 | -0.18 | |
| 同心假浮萤丰度 | 0.25* | 0.09 | 0.03 | -0.17 | |
| 其他种类丰度 | 0.14 | -0.02 | 0.41** | -0.05 |
注: *表示p<0.05, 相关性显著; **表示p<0.01, 相关性极显著 |
| 1 |
陈瑞祥, 林景宏, 曾文阳 , 1992. 黑潮源地与台湾海峡西侧浮游介形类的分布[J]. 海洋学报, 14(5):101-106 (in Chinese).
|
| 2 |
陈瑞祥, 林景宏 , 1993 a. 南黄海和东海浮游介形类的生态特征[J]. 海洋学报, 15(5):104-111 (in Chinese).
|
| 3 |
陈瑞祥, 林景宏 , 1993 b. 南海中部海域浮游介形类的生态研究[J]. 海洋学报, 15(6):91-98 (in Chinese).
|
| 4 |
管秉贤, 袁耀初 , 2006. 中国近海及其附近海域若干涡旋研究综述: I. 南海和台湾以东海域[J]. 海洋学报, 28(3):1-16.
GUAN BINGXIAN, YUAN YAOCHU, 2006. Overview of studies on some eddies in the China seas and their adjacent seas: I. The South China Sea and the region East of Taiwan[J]. Acta Oceanologica Sinica, 28(3):1-16 (in Chinese with English abstract).
|
| 5 |
郭伟龙, 王亮根, 杜飞雁 , 等, 2015. 南沙群岛海域春、秋季浮游介形类生态学特征[J]. 水产学报, 39(6):836-845.
GUO WEILONG, WANG LIANGGEN, DU FEIYAN, et al, 2015. Ecological characteristics of planktonic ostracods in the Nansha Islands waters in spring and autumn[J]. Journal of Fisheries of China, 39(6):836-845 (in Chinese with English abstract).
|
| 6 |
韩舞鹰, 王明彪, 马克美 , 1990. 我国夏季最低表层水温海区—琼东沿岸上升流区的研究[J]. 海洋与湖沼, 21(3):267-275.
HAN WUYING, WANG MINGBIAO, MA KEMEI, 1990. On the lowest surface water temperature area of China sea in summer—the upwelling along the east coast of Hainan Island[J]. Oceanologia et Limnologia Sinica, 21(3):267-275 (in Chinese with English abstract).
|
| 7 |
连喜平, 谭烨辉, 刘永宏 , 等, 2013. 两种浮游生物网对南海北部浮游动物捕获效率的比较[J]. 热带海洋学报, 32(3):33-39.
LIAN XIPING, TAN YEHUI, LIU YONGHONG, et al, 2013. Comparison of capture efficiency for zooplankton in the northern South China Sea, using two plankton mesh sizes[J]. Journal of Tropical Oceanography, 32(3):33-39 (in Chinese with English abstract).
|
| 8 |
林景宏, 陈明达, 陈瑞祥 , 1996. 东海浮游介形类对海流、水团的指示作用[J]. 海洋学报, 18(3):86-91 (in Chinese).
|
| 9 |
徐兆礼, 陈亚瞿 , 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4):13-15.
XU ZHAOLI, CHEN YAQU, 1989. Aggregated intensity of dominant species of zooplankton in autumn in the East China Sea and Yellow Sea[J]. Journal of Ecology, 8(4):13-15 (in Chinese with English abstract).
|
| 10 |
徐兆礼 , 2006. 东海浮游介形类(Ostracods)分布特征[J]. 海洋学报, 28(2):101-108.
XU ZHAOLI, 2006. Characteristics for distribution of pelagic Ostracods in the East China Sea[J]. Acta Oceanologica Sinica, 28(2):101-108 (in Chinese with English abstract).
|
| 11 |
尹健强, 陈清潮 , 1991a. 南沙群岛及其邻近海区浮游介形类的种类、动物区系和动物地理[M] //中国科学院南沙综合科学考察队. 南沙群岛海区海洋动物区系和动物地理研究专集. 北京: 海洋出版社: 64-139(in Chinese).
|
| 12 |
尹健强, 陈清潮 , 1991b. 南沙群岛海区的浮游介形类(1984—1988)[M] //中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物研究论文集(二). 北京: 海洋出版社: 134-154(in Chinese).
|
| 13 |
赵汉取, 李纯厚, 杜飞雁 , 等, 2007. 北部湾海域浮游介形类物种组成、丰度分布及多样性[J]. 生态学报, 27(1):25-33.
ZHAO HANQU, LI CHUNHOU, DU FEIYAN, et al, 2007. Species composition, abundance distribution and diversity of Planktonic Ostracoda in the Beibu Gulf[J]. Acta Ecologica Sinica, 27(1):25-33 (in Chinese with English abstract).
|
| 14 |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.6-2007 海洋调查规范第6部分: 海洋生物调查[S]. 北京: 中国标准出版社.
GENERAL ADMINISTRATION OF QUALITY SUPERVISION, INSPECTION AND QUARANTINE OF THE PEOPLE’S REPUBLIC OF CHINA, STANDARDIZATION ADMINISTRATION OF THE PEOPLE’S REPUBLIC OF CHINA, 2008. GB/T 12763.6-2007 Specifications for oceanographic survey—Part 6: marine biological survey[S]. Beijing: China Standard Publishing House (in Chinese).
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}