
微量元素锰对底栖甲藻热带库里亚藻(Coolia tropicalis)生长及叶绿素荧光特性的影响
|
刘莎莎(1995—), 女, 安徽省亳州市人, 硕士研究生, 研究方向为藻类光合生理, E-mail:shasha_icon@163.com |
收稿日期: 2020-01-08
要求修回日期: 2020-01-29
网络出版日期: 2020-05-19
基金资助
国家自然科学基金(41706126)
国家自然科学基金(41876173)
科技基础资源调查专项(2018FY100200)
科技基础资源调查专项(2018FY100100)
版权
Effect of manganese on the growth and chlorophyll fluorescence characteristics of benthic dinoflagellate Coolia tropicalis
Received date: 2020-01-08
Request revised date: 2020-01-29
Online published: 2020-05-19
Supported by
Foundation item: National Natural Science Fund ation of China(41706126)
National Natural Science Fund ation of China(41876173)
Science & Technology Basic Resources Investigation Program of China(2018FY100200)
Science & Technology Basic Resources Investigation Program of China(2018FY100100)
Copyright
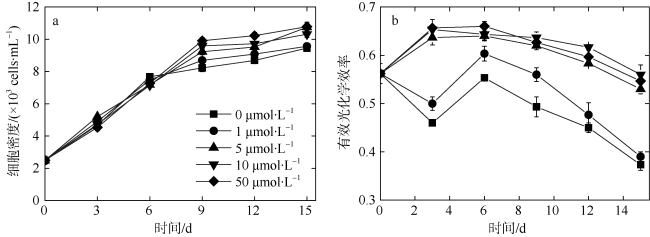
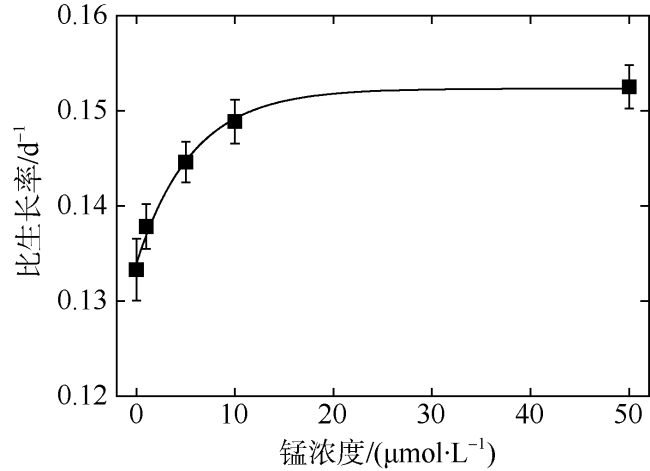
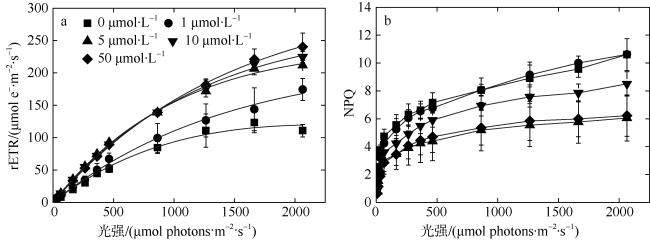
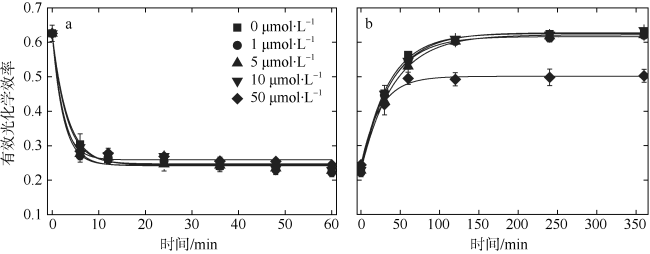
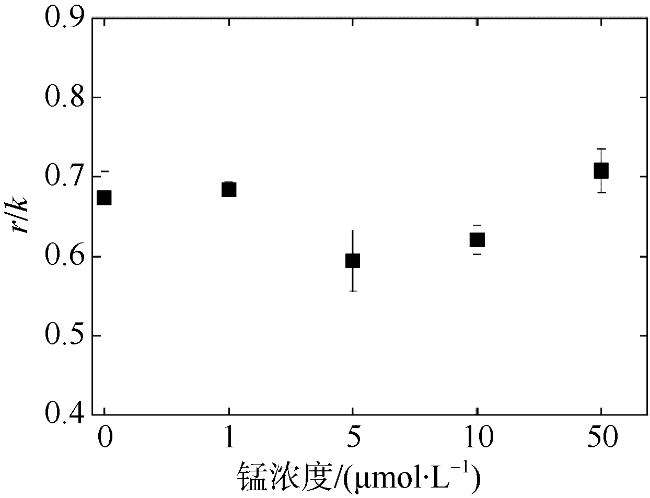
以热带库里亚藻(Coolia tropicalis)为研究对象, 在不同锰浓度(0、1、5、10、50μmol·L -1)的人工海水培养15d, 利用叶绿素荧光动力学技术研究了其生长和光合作用对不同锰环境的响应。结果表明: 1)比生长速率(μ)和最大相对电子传递速率(rETRmax)与锰浓度均呈指数关系且对锰胁迫具有相同程度的响应; 2)锰浓度至少大于1μmol·L -1才能维持热带库里亚藻正常的光合作用活性, 当锰浓度低于该浓度时, 光合作用活性(Fv/Fm)在6d后开始下降, 而单位反应中心吸收光能(ABS/RC)和热能耗散(DI0/RC)升高; 两个反应中心之间的电子传递(φE0)及生长并未受影响, 表明此阶段锰缺乏只影响活性光反应中心数量并提高热耗散途径; 当锰缺乏延长至15d时, 胁迫作用显现(F0上升)并且电子传递(φE0)和生长受到抑制, 这阶段锰缺乏使光反应中心关闭且电子传递受阻; 3)锰缺乏的修复损伤比(r/k)并未降低, 表明锰缺乏并未影响热带库里亚藻的光保护能力。
刘莎莎 , 陈亨 , 黄凯旋 , 吕颂辉 , 张亮 , 谢学东 . 微量元素锰对底栖甲藻热带库里亚藻(Coolia tropicalis)生长及叶绿素荧光特性的影响[J]. 热带海洋学报, 2020 , 39(3) : 76 -85 . DOI: 10.11978/2020003
In this study, the effects of growth and chlorophyll fluorescence characteristics in Coolia tropicalis under different concentrations of manganese (Mn, 0, 1, 5, 10, and 50μmol·L -1) were investigated by using chlorophyll fluorescence kinetics technique. The results are as follows. The relationships of specific growth rate (μ) and maximal relative electron transport rate (rETRmax) with the concentration of Mn were fitted by exponential functions; growth and electron transport had the same degree of response to manganese stress. The regular activity of photosynthesis can be maintained only when the concentration of manganese is at least 1μmol·L -1. when the Mn concentration was lower than that 1μmol·L -1, the photochemical efficiency (Fv/Fm) begins to decrease after 6 days, while the per reaction center of absorbed light energy (ABS/RC) and heat energy dissipation (DI0/RC) increase, and no influence to electron transport (φE0) and the growth, which indicated Mn deficiency only decreased the number of active reaction center and increased the heat dissipation in this phase. While Mn deficiency extended to day 15, the rising of F0 and the decreases of φE0 and growth indicated that Mn deficiency caused irreversible damage to the active reaction center and inhibited the electron transport in this phase. The repair rate to damage rate (r/k) under intense light was not affected by Mn deficiency, indicated Mn deficiency did not influence the photoprotection of C. tropicalis.
表1 JIP-测定的相关参数Tab. 1 The coefficients of JIP-test |
| 参数及其计算 | 定义 |
|---|---|
| F0 | 暗适应后的最小荧光强度 |
| Fm=FP | 暗适应后的最大荧光值 |
| VJ = (FJ - F0) / (Fm - F0) | 在J点的相对可变荧光 |
| VI = (FI - F0) / (Fm - F0) | 在I点的相对可变荧光 |
| Fv / F0 = (Fm - F0) -1 | PSⅡ的潜在活性 |
| Fv / Fm | PSⅡ最大光化学效率 |
| M0 = 4 (F300μs - F0) / (Fm - F0) | OJIP荧光诱导曲线的初始斜率 |
| Area | 荧光诱导曲线与F=Fm之间的面积 |
| Sm = Area / (Fm - F0) | 标准化后的在OJIP荧光诱导曲线和F=Fm之间的面积 |
| ABS / RC = M0 (1/VJ) · (1/φP0) | 单位反应中心吸收的光能 |
| TR0 / RC = M0 (1/VJ) | 单位反应中心捕获的用于还原QA的能量(在t=0时) |
| ET0 / RC = M0 (1/VJ)ψ0 | 单位反应中心捕获的用于电子传递的能量(在t=0时) |
| DI0 / RC = (ABS / RC) - (TR0 / RC) | 单位反应中心的热耗散(在t=0时) |
| φP0 = TR0 / ABS =(1- F0/Fm) | 最大光化学效率(在 t=0 时) |
| φ0 = ET0 / TR0 =(1- VJ) | 捕获的激子将电子传递到电子传递链中超过 QA 的其它电子受体的概率(在t=0时) |
| φE0 = ET0 / ABS =(1- F0 / Fm ) φ0 | 用于电子传递的量子产额(在t=0时) |
| φD0 = 1- φP0 = (F0 / Fm ) | 用于热耗散的量子比率(在t=0时) |
| PIABS = RC/ABS×[φP0 /(1- φP0 )] [φ0/ (1- φ0 )] | 以吸收光能为基础的性能指数 |
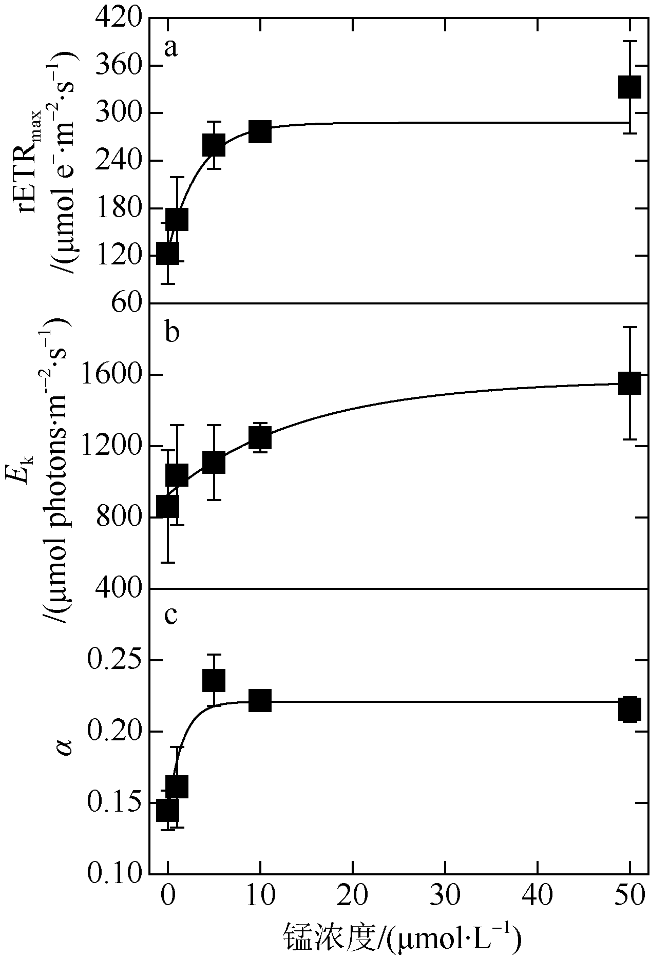
表2 μ, rETRmax, Ek和α拟合曲线的初始斜率及各参数下的饱和锰浓度Tab. 2 Initial slopes and saturated manganese concentrations of μ, rETRmax, Ek, and α |
| 参数 | 初始斜率 | 饱和锰浓度/(μmol·L-1) |
|---|---|---|
| μ/(d-1) | 0.24 ± 0.03 | 48.66 ± 5.49 |
| rETRmax/(μmol e-·m-2·s-1) | 0.25 ± 0.02 | 17.29 ± 1.12 |
| Ek/(μmol photons·m-2·s-1) | 0.09 ± 0.04 | 49.74 ± 19.67 |
| α | 0.31 ± 0.03 | 15.72 ± 1.47 |
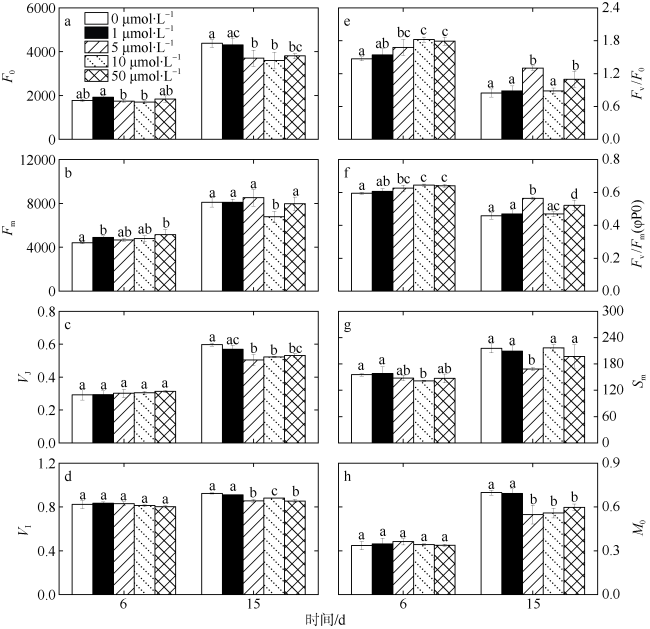
图5 第6和15d不同浓度锰处理下热带库里亚藻的叶绿素荧光参数变化a.最小荧光值; b.最大荧光值; c. 在 J 点的相对可变荧光强度; d. 在 I 点的相对可变荧光强度; e. PSⅡ 潜在活性; f. 最大光化学效率; g. 标准化后的在 OJIP 荧光诱导曲线和 F=Fm 之间的面积; h. OJIP 荧光诱导曲线的初始斜率; 图中不同字母表示有显著性差异 Fig. 5 Changes of chlorophyll fluorescence induction parameters of C. tropicalis under different Mn treatment on days 6 and 15. a) minimal fluorescence intensity; b) maximal fluorescence intensity; c) relative variable fluorescence intensity at the J-step; d) relative variable fluorescence intensity at the I-step; e) potential activity of PSⅡ; f) maximal photochemical efficiency; g) normalised total complementary area above the O-J-I-P transie; h) approximated initial slope of the fluorescence transient |
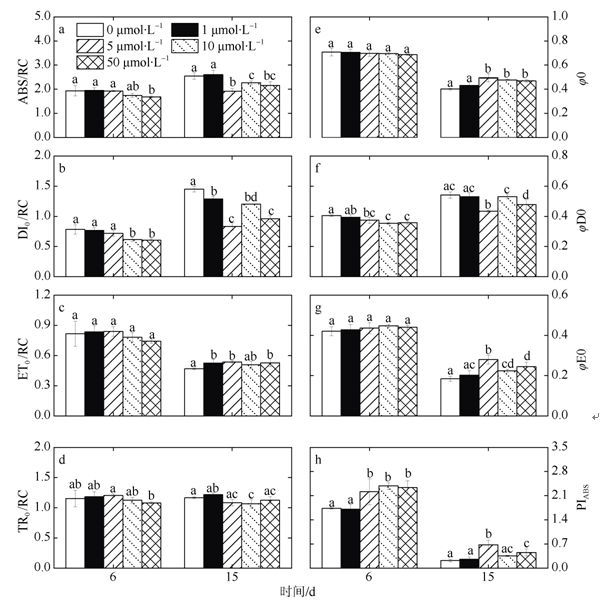
图6 第6和15d不同浓度锰处理下ABS/RC、DI0/RC、ET0/RC、TR0/RC、φ0、φD0、φE0和 PIABS变化a. 单位反应中心吸收的光能; b. 单位反应中心的热耗散; c. 单位反应中心捕获的用于电子传递的能量; d. 单位反应中心捕获的用于还原 QA 的能量; e. 捕获的激子将电子传递到电子传递链中超过 QA 的其它电子受体的概率; f. 用于热耗散的量子比率; g. 用于电子传递的量子产额; h. 以吸收光能为基础的性能指数; 图中不同字母表示有显著性差异 Fig. 6 Changes of ABS/RC, DI0/RC, ET0/RC, TR0/RC, φ0, φD0, φE0, and PIABS under different Mn treatment on days 6 and 15. a) absorption flux per reaction center; b) dissipated energy flux per reaction center; c) electron transport flux per reaction center; d) trapped energy flux per reaction center; e) probability that a trapped exciton moves an electron into the electron transport chain beyond QA-; f) quantum yield for energy dissipation; g) quantum yield for electron transport; h) performance index on absorption basis |
| 1 |
曹春晖, 孙世春, 王学魁 , 等, 2010. 锰浓度对米氏凯伦藻叶绿素荧光特性及生长的影响[J]. 生态学报, 30(19):5280-5288.
|
| 2 |
郭皓, 丁德文, 林凤翱 , 等, 2015. 近20a我国近海赤潮特点与发生规律[J]. 海洋科学进展, 33(4):547-558.
|
| 3 |
梁计林, 龙丽娟, 张偲 , 等, 2011. 微量元素Fe、Mn、Co对有毒甲藻生长和产毒的影响[J]. 热带海洋学报, 30(1):119-123.
|
| 4 |
梁舜华, 张红标 , 1993. 大鹏湾盐田水域赤潮期间水质锰的变化规律[J]. 海洋通报, 12(2):13-16.
|
| 5 |
沈竑, 洪君超, 张开富 , 等, 1995. 中肋骨条藻(Skeletonema costatum)赤潮发生过程中微量元素Fe、Mn作用的研究[J]. 暨南大学学报(自然科学与医学版), 16(1):131-136, 149.
|
| 6 |
王木兰, 姜玥璐 , 2018. 微量元素锰对威氏海链藻生长及叶绿素荧光的影响[J]. 环境科学, 39(12):5514-5522.
|
| 7 |
杨秀环, 唐宝英, 吴京洪 , 等, 2000. 柘林湾赤潮与Fe、Mn、Se和营养盐指数的关系[J]. 中山大学学报(自然科学版), 39(5):58-62.
|
| 8 |
叶波, 吴永波, 邵维 , 等, 2014. 高温干旱复合胁迫及复水对构树(Broussonetia papyrifera)幼苗光合特性和叶绿素荧光参数的影响[J]. 生态学杂志, 33(9):2343-2349.
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}