
中国西沙群岛底栖甲藻热带库里亚藻(Coolia tropicalis)的形态学、系统发育及毒性研究
|
黄丽芬(1995—), 女, 广东省清远市人, 硕士研究生, 研究方向为藻类分类。E-mail:hhhliii22@163.com |
收稿日期: 2019-12-11
要求修回日期: 2020-02-03
网络出版日期: 2020-05-19
基金资助
国家自然科学基金项目(41876173)
国家自然科学基金项目(41706126)
科技基础资源调查专项(2018FY100200)
科技基础资源调查专项(2018FY100200、2018FY100100)(2018FY100100)
版权
Morphology, phylogeny and toxicity of Coolia tropicalis (Dinophyceae) from the Xisha Islands, China
Received date: 2019-12-11
Request revised date: 2020-02-03
Online published: 2020-05-19
Supported by
Foundation item: National Natural Science Foundation of China(41876173)
Foundation item: National Natural Science Foundation of China(41706126)
Science & Technology Basic Resources Investigation Program of China(2018FY100200)
Science & Technology Basic Resources Investigation Program of China(2018FY100100)
Copyright
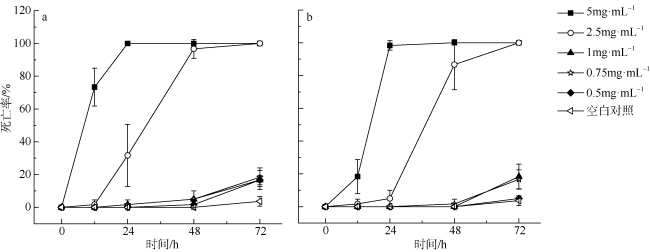
库里亚藻(Coolia)是一类广泛分布且具有生物毒性的海洋底栖甲藻。本研究对近年来在中国南海西沙群岛海域采集分离的4株库里亚藻, 通过光镜及电镜下的形态学观察, 并结合基于核糖体大亚基(large subunit, LSU)rDNA(D1-D3)和内部转录间隔区(internal transcribed spacer, ITS)的序列进行系统发育分析, 鉴定该4个株系为热带库里亚藻(Coolia tropicalis)。在卤虫(Artemia salina)生物毒性试验中, 热带库里亚藻不同株系毒素提取液对卤虫幼虫表现出了毒性差异, XS554株系的48h半致死浓度(LC50)为1.42mg·mL -1(约相当于1.1×10 5cells·mL -1), 5XS15株系的LC50为1.92mg·mL -1(约相当于1×10 5cells·mL -1)。
黄丽芬 , 李群 , 吕颂辉 , 张亮 , 谢学东 . 中国西沙群岛底栖甲藻热带库里亚藻(Coolia tropicalis)的形态学、系统发育及毒性研究[J]. 热带海洋学报, 2020 , 39(3) : 86 -97 . DOI: 10.11978/2019129
Coolia is a widespread and toxic genus of benthic marine dinoflagellate. In this study, four strains of Coolia species were examined as Coolia tropicalis, which were collected in the Xisha Islands of the South China Sea in recent years. Morphology was determined using light microscopy and scanning electron microscopy. Phylogenetic analyses were constructed based on the sequences of the large subunit rDNA and the internal transcribed spacer region. The bioassay toxicity test of brine shrimp (Artemia salina) demonstrated that the algal lysates extracted from different strains of C. tropicalis exhibited different toxic effects. At 48 h, the LC50 of XS554 strain was 1.42 mg·mL -1 (approximately 1.1×10 5 cells·mL -1) and 5XS15 strain was 1.92 mg·mL -1 (approximately 1×10 5 cells·mL -1).
Key words: Coolia tropicalis; morphology; phylogeny; toxicity; Artemia salina
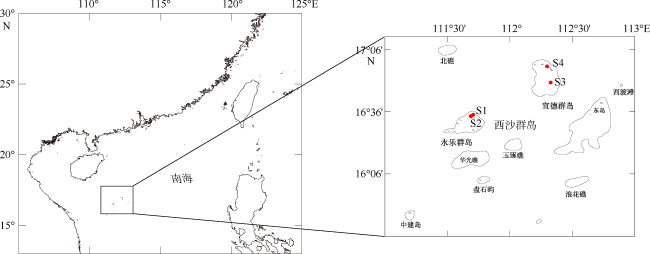
表1 藻株采样信息Tab. 1 Sampling information |
| 株系号 | 采样时间 | 采样点 | 经纬度 | 附着基质 |
|---|---|---|---|---|
| 2XS73 | 2016-06-11 | 银屿岛(S1) | 16°34'15.39"N, 111°42'13.71"E | 死珊瑚, 少量活珊瑚 |
| 3XS45 | 2016-06-12 | 鸭公岛(S2) | 16°33'30.29"N, 111°40'40.56"E | 死珊瑚 |
| XS554 | 2015-06-11 | 永兴岛(S3) | 16°33'30.29"N, 111°40'40.56"E | 礁岩底质 |
| 5XS15 | 2015-06-11 | 赵述岛(S4) | 16°57'18.73"N, 112°15'58.50"E | 礁石底, 死珊瑚 |
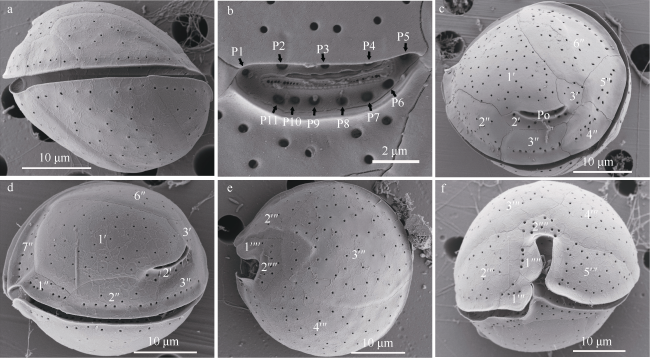
图3 热带库里亚藻的扫描电镜图a. 侧面观; b. 顶孔复合体; c. 顶面观; d. 顶面观; e. 底面观; f. 腹面观 Fig. 3 Scanning electron microscope (SEM) images of Coolia tropicalis. a) Lateral view; b) close-up of APC; c) apical view of the epitheca; d) apical view of the epitheca; e) antapical view of the hypotheca; f) ventral view |
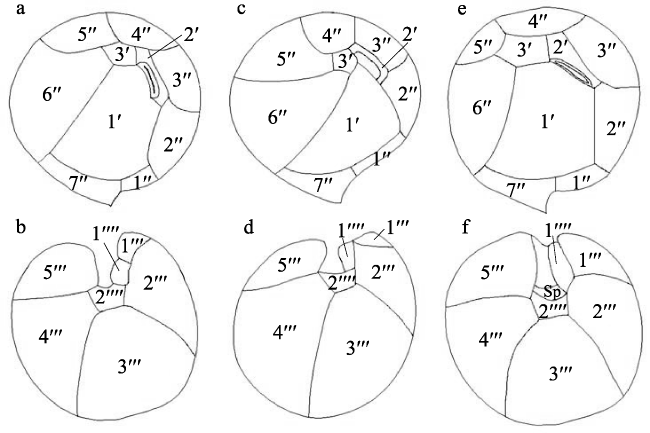
图4 热带库里亚藻的壳板图a、b, 本研究; c、d, 重绘自Faust(1995); e、f, 重绘自。a、c、e为上壳板顶面观; b、d、f为下壳板底面观 Fig. 4 Thecal plate tabulation of C. tropicalis based on original descriptions. (a, b) This study; (c, d) redrawn from Faust(1995); (e, f) redrawn from Mohammad-Noor et al (2013); (a, c, e) apical view of the epitheca; (b, d, f) antapical view of the hypotheca |
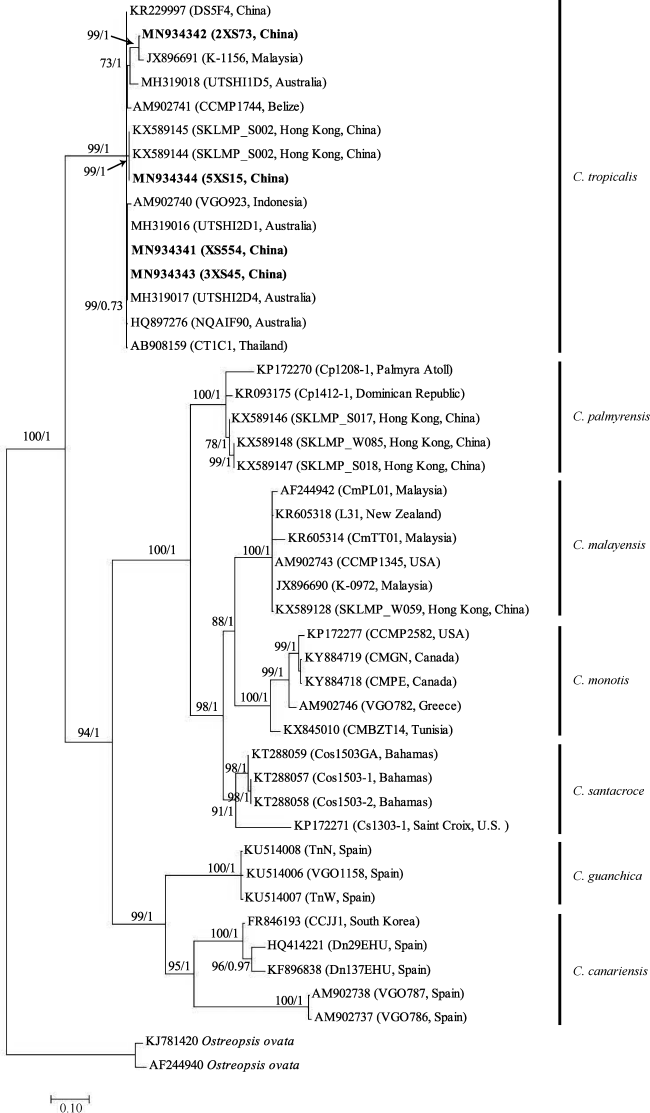
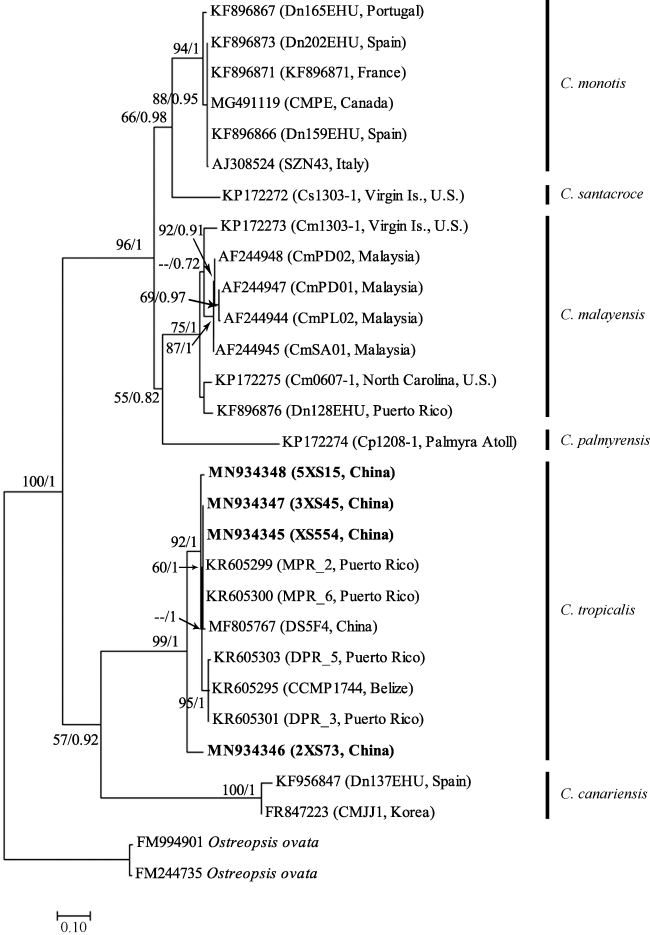
图5 基于核糖体大亚基(LSU)D1-D3区序列的系统发育树以Ostreopsis ovata作为外类群, 分支节点处的数值表示对该分支的统计支持值, 分别为最大似然法自举检验值(100)和贝叶斯推断后验概率值(1) Fig. 5 Molecular phylogenetic tree based on the D1-D3 region of LSU rDNA with Ostreopsis ovata as outgroups. The numbers at nodes indicate the maximum likelihood booststrap (BS) values (100) and Bayesian Inference posterior probability (PP) values (1), representing the levels of support |
表2 库里亚藻不同物种间基于LSU rDNA序列的遗传距离Tab. 2 Genetic distances of Coolia among different species based on LSU rDNA |
| 种名 | Coolia tropicalis | Coolia palmyrensis | Coolia malayensis | Coolia monotis | Coolia santacroce | Coolia guanchica |
|---|---|---|---|---|---|---|
| C. tropicalis | ||||||
| C. palmyrensis | 0.421 | |||||
| C. malayensis | 0.434 | 0.146 | ||||
| C. monotis | 0.450 | 0.170 | 0.117 | |||
| C. santacroce | 0.428 | 0.098 | 0.090 | 0.086 | ||
| C. guanchica | 0.365 | 0.394 | 0.431 | 0.425 | 0.361 | |
| C. canariensis | 0.392 | 0.354 | 0.396 | 0.416 | 0.359 | 0.187 |
图6 基于核糖体转录间隔区ITS序列的系统发育树核糖体以Ostreopsis ovata作为外类群, 分支节点处的数值表示对该分支的统计支持值, 分别为最大似然法自举检验值(100)和贝叶斯推断后验概率值(1) Fig. 6 Molecular phylogenetic tree based on the ITS region with Ostreopsis ovata as outgroups. The numbers at nodes indicate the maximum likelihood booststrap (BS) values (100) and Bayesian Inference posterior probability (PP) values (1), representing the levels of support |
表3 库里亚藻不同物种间基于ITS区序列的遗传距离Tab. 3 Genetic distances of Coolia among different species based on ITS region |
| 种名 | Coolia tropicalis | Coolia canariensis | Coolia palmyrensis | Coolia santacroce | Coolia monotis |
|---|---|---|---|---|---|
| C. tropicalis | |||||
| C. canariensis | 0.552 | ||||
| C. palmyrensis | 0.553 | 0.798 | |||
| C. santacroce | 0.477 | 0.737 | 0.397 | ||
| C. monotis | 0.428 | 0.577 | 0.379 | 0.201 | |
| C. malayensis | 0.539 | 0.651 | 0.379 | 0.209 | 0.128 |
| 1 |
刘宁宁, 林永水 , 1999. 有毒甲藻Coolia monotis Meunier的研究I. 甲板形态[J]. 热带海洋, 18(2):1-4.
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}