
北部湾棕囊藻藻华原因种分析
|
徐轶肖(1977—), 女, 浙江省兰溪市人, 副研究员, 从事海洋有毒有害藻与海产品安全研究。email: |
Copy editor: 林强
收稿日期: 2020-03-16
要求修回日期: 2020-06-15
网络出版日期: 2020-06-15
基金资助
国家自然科学基金(41976155)
国家自然科学基金(41506137)
广西自然科学基金(2016GXNSFBA380037)
版权
Causative species of Phaeocystis blooms in Beibu Gulf
Copy editor: LIN Qiang
Received date: 2020-03-16
Request revised date: 2020-06-15
Online published: 2020-06-15
Supported by
National Natural Science Foundation of China(41976155)
National Natural Science Foundation of China(41506137)
Natural Science Foundation of Guangxi(2016GXNSFBA380037)
Copyright
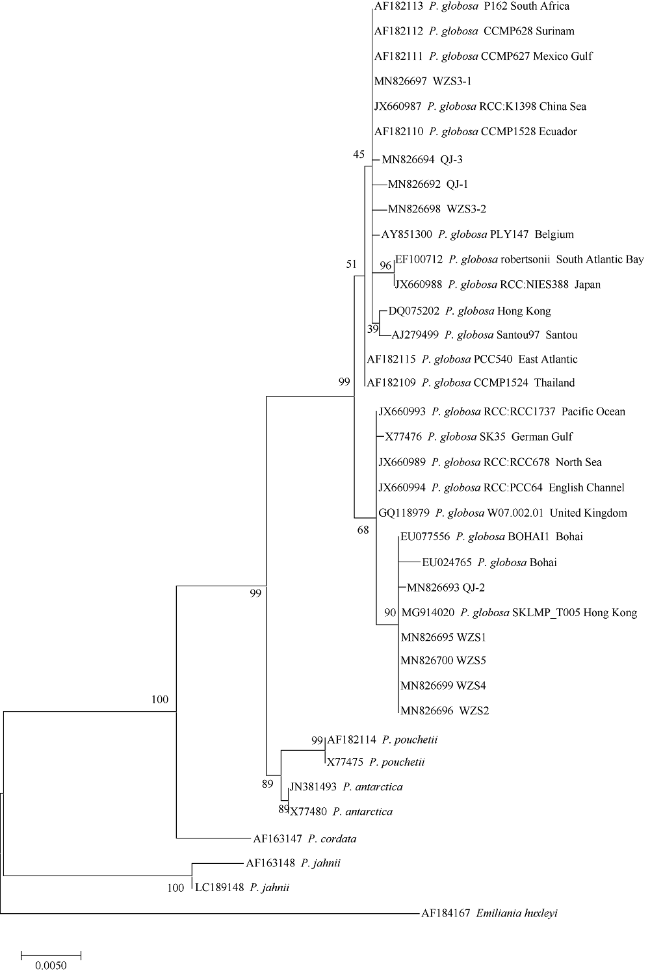
近年来, 棕囊藻已成为我国北部湾海域藻华发生的主要原因种, 严重影响北部湾海洋生态环境、水产增养殖业和防城港核电安全, 急切需要对棕囊藻藻华原因种作进一步分析确认。2017年2月和3月在钦州湾和涠洲岛藻华发生海域分离得到9株棕囊藻, 采用光学显微镜进行形态学初步鉴定, 并通过核糖体小亚基序列(SSU rDNA)进行系统进化分析。研究结果表明, 北部湾9株棕囊藻与球形棕囊藻形态特征基本符合, 系统发育树与不同海域来源的球形棕囊藻聚在同一大分支上, 与其他海域来源的球形棕囊藻遗传距离为0.0000~0.0071, 其中北部湾WZS3-1株与墨西哥湾株CCMP627、苏里南株CCMP628、厄瓜多尔株CCMP1528、南非株P162、中国海株RCC:K1398及汕头株Santou97的球形棕囊藻亲缘关系最近, 遗传距离为0.0000, 而北部湾WZS1、WZS2、WZS4、WZS5株与香港株SKLMP_T005和渤海株BOHAI1亲缘关系最近, 遗传距离为0.0000。球形棕囊藻的种内遗传距离(0.0000~0.0073)明显小于种间遗传距离(0.0084~0.0440), 因此可确定来自北部湾不同海域的9株棕囊藻均为球形棕囊藻。此外, 球形棕囊藻各藻株间的亲缘关系远近与其地理位置分布不能完全对应。本研究结果将为我国棕囊藻藻华藻种库构建、棕囊藻北部湾株生理生态研究和北部湾棕囊藻藻华发生机理研究等提供参考。
徐轶肖 , 何喜林 , 张腾 , 蓝文陆 . 北部湾棕囊藻藻华原因种分析[J]. 热带海洋学报, 2020 , 39(6) : 122 -130 . DOI: 10.11978/2020030
In recent years, Phaeocystis has become the main causative species for algal blooms in the Beibu Gulf of China. It has deleterious effects on marine ecosystem and aquaculture, and causes nuclear power plant safety problems in Fangcheng. It is thus urgent to further analyze the causative species of Phaeocystis in the region. In February and March 2017, nine strains of Phaeocystis sp. identified by optical microscope were isolated during algal blooms in Qinzhou Bay and Weizhou Island; their SSU rDNA sequences were then analyzed using Maximum Likelihood (ML) phylogenetic tree. The results showed that all nine strains of Phaeocystis sp. were morphologically consistent with those of Phaeocystis globosa. Their genetic distance varied between 0.0000-0.0073; and in the phylogenetic tree, all clustered in the same branch with those P. globosa from other seawaters. Among them, WZS3-1 strain in the Beibu Gulf had the closest genetic relationship with P. globosa from Gulf of Mexico CCMP627, Surinam CCMP628, Ecuador CCMP1528, South Africa P162, China Sea RCC:K1398 and Santou97, exhibiting a genetic distance of 0.0000. Strains of WZS1, WZS2, WZS4, and WZS5 in the Beibu Gulf had a genetic distance of 0.0000 with Hong Kong SKLMP_T005 and Bohai BOHAI1. The intraspecific genetic distance of P. globosa (0.0000-0.0071) is significantly smaller than the interspecific genetic distance of P. globosa (0.0084-0.0440). As a result, these nine strains from Beibu Gulf all belong to P. globosa. The phylogenetic distance for strains of P. globosa do not reflect their geographical distance. The findings of this study are essential for Phaeocystis culture collection, their further ecophysiological study and understanding of occurrence mechanisms for Phaeocystis blooms in the Beibu Gulf.
表1 样品信息Tab. 1 Information on sampling sites |
| 株名 | 采集地 | 采集时间 |
|---|---|---|
| QJ-1 | 钦州湾(21°41′38″N, 108°35′13″E) | 2017-02-22 |
| QJ-2 | 钦州湾(21°41′38″N, 108°35′13″E) | 2017-02-22 |
| QJ-3 | 钦州湾(21°41′38″N, 108°35′13″E) | 2017-02-22 |
| WZS1 | 涠洲岛(21°03′16″N, 109°08′71″E) | 2017-03-21 |
| WZS2 | 涠洲岛(21°06′44″N, 109°10′36″E) | 2017-03-21 |
| WZS3-1 | 涠洲岛(21°06′48″N, 109°13′43″E) | 2017-03-21 |
| WZS3-2 | 涠洲岛(21°06′48″N, 109°13′43″E) | 2017-03-21 |
| WZS4 | 涠洲岛(21°03′23″N, 109°13′15″E) | 2017-03-21 |
| WZS5 | 涠洲岛(21°02′42″N, 109°11′13″E) | 2017-03-21 |
表2 棕囊藻SSU rDNA基因序列Tab. 2 SSU rDNA sequences of Phaeocystis spp. |
| 种 | 株系 | 来源地 | 登录号 |
|---|---|---|---|
| P. globosa | QJ-1 | 中国, 钦州湾 | MN826692 |
| P. globosa | QJ-2 | 中国, 钦州湾 | MN826693 |
| P. globosa | QJ-3 | 中国, 钦州湾 | MN826694 |
| P. globosa | WZS1 | 中国, 涠洲岛 | MN826695 |
| P. globosa | WZS2 | 中国, 涠洲岛 | MN826696 |
| P. globosa | WZS3-1 | 中国, 涠洲岛 | MN826697 |
| P. globosa | WZS3-2 | 中国, 涠洲岛 | MN826698 |
| P. globosa | WZS4 | 中国, 涠洲岛 | MN826699 |
| P. globosa | WZS5 | 中国, 涠洲岛 | MN826670 |
| P. globosa | - | 中国香港 | DQ075202 |
| P. globosa | SKLMP_T005 | 中国香港 | MG914020 |
| P. globosa | BOHAI1 | 中国, 渤海 | EU077556 |
| P. globosa | - | 中国, 渤海 | EU024765 |
| P. globosa | RCC:K1398 | 太平洋-中国海域 | JX660987 |
| P. globosa | Santou97 | 中国, 汕头 | AJ279499 |
| P. globosa | CCMP628 | 大西洋, 苏里南 | AF182112 |
| P. globosa | W07.002.01 | 英国 | GQ118979 |
| P. globosa | robertsonii | 南大西洋湾 | EF100712 |
| P. globosa | CCMP627 | 墨西哥湾 | AF182111 |
| P. globosa | SK35 | 德国湾 | X77476 |
| P. globosa | PLY147 | 比利时 | AY851300 |
| 种 | 株系 | 来源地 | 登录号 |
| P. globosa | CCMP1528 | 厄瓜多尔 | AF182110 |
| P. globosa | P162 | 南非 | AF182113 |
| P. globosa | CCMP1524 | 泰国 | AF182109 |
| P. globosa | RCC:PCC64 | 英吉利海峡 | JX660994 |
| P. globosa | RCC:RCC678 | 英国, 北海(大西洋) | JX660989 |
| P. globosa | RCC:NIES388 | 日本 | JX660988 |
| P. globosa | PCC540 | 东大西洋 | AF182115 |
| P. globosa | RCC:RCC1737 | 太平洋 | JX660993 |
| P. pouchetii | P360 | 挪威, 劳内峡湾 | AF182114 |
| P. pouchetii | SK34 | 格陵兰岛 | X77475 |
| P. antarctica | SK22 | 南大洋威德尔海 | X77480 |
| P. antarctica | MM-E1B6 | - | JN381493 |
| P. jahnii | NIES-3884 | 地中海 | LC189148 |
| P. jahnii | CCMP2496 | 地中海 | AF163148 |
| P. cordata | - | 地中海 | AF163147 |
| Emiliania huxleyi | Texel B | - | AF184167 |
图1 北部湾球形棕囊藻 (WZS1株)a、b. 具鞭毛游动单细胞, a中箭头指鞭毛, b中箭头指叶绿体; c、d. 不具鞭毛的不动单细胞, d中箭头指细胞核; e. 群体/囊体; f. 野外现场样品; g. 藻华现场海水; h. 藻华消退产生的白色泡沫 Fig. 1 Phaeocystis globosa in Beibu Gulf. (a, b) Swimming single cell, arrows in a and b representing flagella and chloroplast, respectively; (c, d) immobile single cell without flagella, arrow in d showing the nucleus; e) colony; f) field sample; g) bloom seawater; h) white foam produced by Phaeocystis bloom during its degradation stage |
| [1] |
陈菊芳, 徐宁, 江天久, 等, 1999. 中国赤潮新记录种─球形棕囊藻(Phaeocystis globosa)[J]. 暨南大学学报(自然科学版), 20(3):124-129.
|
| [2] |
贺立燕, 宋秀贤, 於凡, 等, 2019. 潜在影响防城港核电冷源系统的藻类暴发特点及其监测防控技术[J]. 海洋与湖沼, 50(3):700-706.
|
| [3] |
胡晓坤, 张清春, 陈振帆, 等, 2019. 北部湾海域球形棕囊藻遗传多样性分析[J]. 海洋与湖沼, 50(3):601-610.
|
| [4] |
胡章喜, 邓蕴彦, 唐赢中, 2019. 我国北部湾球形棕囊藻(Phaeocystis globosa)的表面形态和细胞超微结构的电镜观察[J]. 海洋与湖沼, 50(3):621-629.
|
| [5] |
李亚男, 沈萍萍, 黄良民, 等, 2012. 棕囊藻的分类及系统进化研究进展[J]. 生态学杂志, 31(3):745-754.
|
| [6] |
刘国强, 史海燕, 魏春雷, 等, 2008. 广西涠洲岛海域浮游植物和赤潮生物种类组成的初步研究[J]. 海洋通报, 27(3):43-48.
|
| [7] |
罗金福, 李天深, 蓝文陆, 2016. 北部湾海域赤潮演变趋势及防控思路[J]. 环境保护, 44(20):40-42.
|
| [8] |
覃仙玲, 赖俊翔, 陈波, 等, 2016. 棕囊藻北部湾株的18S rDNA分子鉴定[J]. 热带亚热带植物学报, 24(2):176-181.
|
| [9] |
曲凌云, 吕颂辉, 高春蕾, 等, 2008. 棕囊藻渤海株核糖体18S rDNA和ITS基因结构序列分析[J]. 海洋科学进展, 26(2):200-206.
|
| [10] |
沈萍萍, 齐雨藻, 欧林坚, 2018. 中国沿海球形棕囊藻(Phaeocystis globosa)的分类、分布及其藻华[J]. 海洋科学, 42(10):146-162.
|
| [11] |
侍茂崇, 2014. 北部湾环流研究述评[J]. 广西科学, 21(4):313-324.
|
| [12] |
王锦秀, 孔凡洲, 陈振帆, 等, 2019. 六株球形棕囊藻的色素组成特征研究[J]. 海洋与湖沼, 50(3):611-620.
|
| [13] |
杨士瑛, 陈波, 李培良, 2006. 用温盐资料研究夏季南海水通过琼州海峡进入北部湾的特征[J]. 海洋湖沼通报, ( 1):1-7.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}