
基于线粒体控制区序列的花斑蛇鲻遗传多态性分析
|
李敏(1984—), 男, 湖南长沙人, 副研究员, 博士, 从事海洋生物多样性保护研究。E-mail: |
Copy editor: 姚衍桃
收稿日期: 2018-11-13
要求修回日期: 2020-01-11
网络出版日期: 2020-07-27
基金资助
广东省自然科学基金项目(2014A030310177)
农业农村部财政专项(NFZX2018)
广东省促进海洋经济发展专项资金(GDME-2018E004)
中国水产科学研究院南海水产研究所中央级公益性科研院所基本科研业务费专项资金(2019TS13)
版权
Genetic polymorphism of the Brushtooth lizardfish Saurida undosquamis based on mitochondrial D-loop sequences
Received date: 2018-11-13
Request revised date: 2020-01-11
Online published: 2020-07-27
Supported by
Foundation item: Natural Science Foundation of Guangdong Province(2014A030310177)
Financial Fund of the Ministry of Agriculture and Rural Affairs(NFZX2018)
Special Fund for Promoting Marine Economic Development of Guangdong Province(GDME-2018E004)
Central Public-interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS(2019TS13)
Copyright
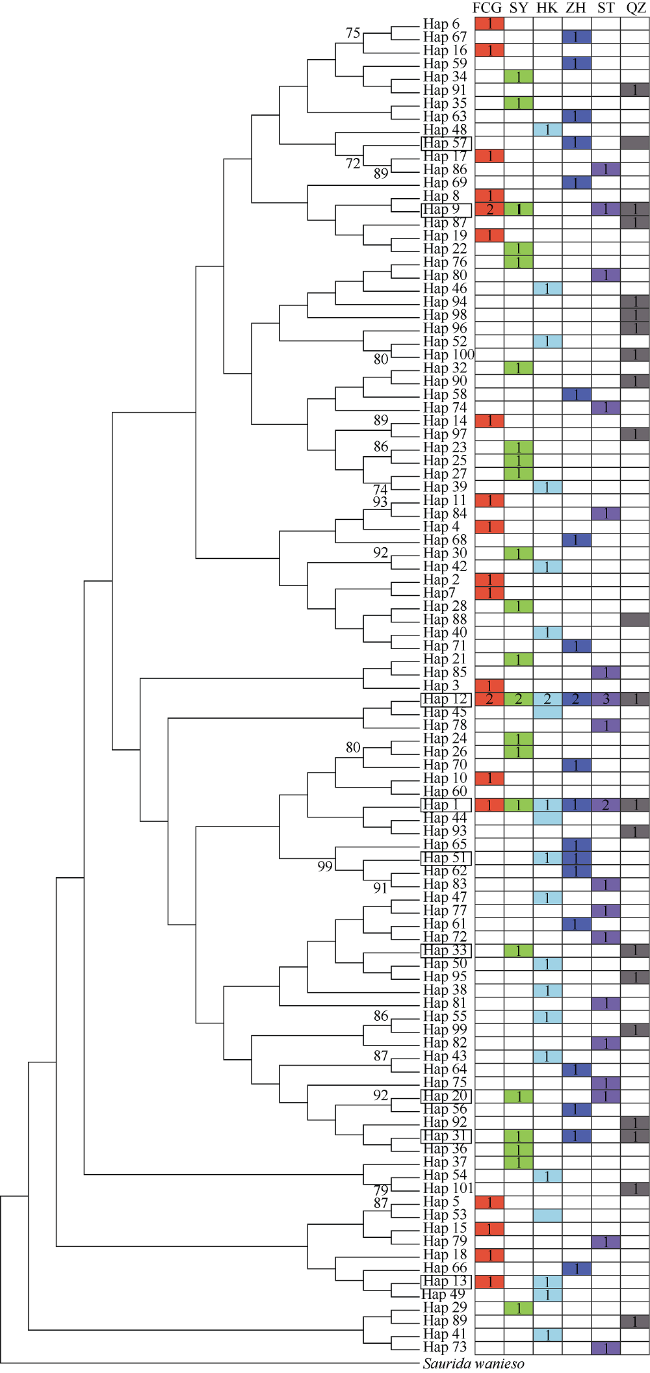
花斑蛇鲻(Saurida undosquamis)是一种重要的底层经济鱼类, 本研究利用线粒体控制区(D-loop区)序列分析了中国近海分布的花斑蛇鲻的遗传结构和遗传多样性, 一共测定了6个地理群体129尾样本的D-loop区全序列。结果显示, 全长为921bp的序列包含71个多态性位点, 共检测到101个单倍型。花斑蛇鲻总体呈现很高的单倍型多样性(0.9873 ± 0.0048)和较低的核苷酸多样性(0.0132 ± 0.0067)的特征。基于邻接法构建的单倍型系统发育树的拓扑结构很浅, 没有形成分化明显的支系。单倍型在各个地理群体中的分布呈分散交叉状态, 表明地理群体间没有显著的遗传分化。6个群体的分子方差分析显示, 花斑蛇鲻绝大部分的遗传变异(99.87%)来源于群体内的个体之间, 而群体间的变异仅占0.13%。大部分地理群体之间的遗传分化指数(FST)均很小, 揭示了群体间的基因交流很频繁, 存在高度的遗传同质性。这些研究结果表明中国近海的花斑蛇鲻遗传多样性丰富, 没有明显的遗传分化, 是一个随机交配群, 可以作为同一个渔业单元来管理。
李敏 , 孔啸兰 , 许友伟 , 陈作志 . 基于线粒体控制区序列的花斑蛇鲻遗传多态性分析[J]. 热带海洋学报, 2020 , 39(4) : 42 -49 . DOI: 10.11978/2019115
The Brushtooth lizardfish (Saurida undosquamis) is one of the economically important demersal fishes. The population genetic structure and genetic diversity of S. undosquamis from the coast of China were examined based on the complete control region (D-loop) sequences. A total of 129 individuals from six geographic populations were sequenced. Seventy-one polymorphic sites were detected, which defined 101 haplotypes. Results show S. undosquamis from the coast of China are characterized by quite high haplotype diversity (0.9873 ± 0.0048) and relative low nucleotide diversity (0.0132 ± 0.0067). Phylogenetic tree for haplotypes based on Neighbour-joining method shows shallow topology and reveals no significant divergent clades. Haplotypes from each geographic population were scattered throughout the NJ tree, showing no significant genetic differentiation between populations. Analyses of molecular variance suggest nearly all the genetic variation (99.87%) is attributed to variability within populations, while little variations (0.13%) are found between populations. Most of the pairwise FST values between different populations are quite low, which implies a high rate of gene flow and genetic homogeneity between populations. The results demonstrate high genetic diversity and little genetic differentiation for S. undosquamis from the coast of China. They belong to the same population (panmixia), and a single-stock management regime could be supported in fishery management.
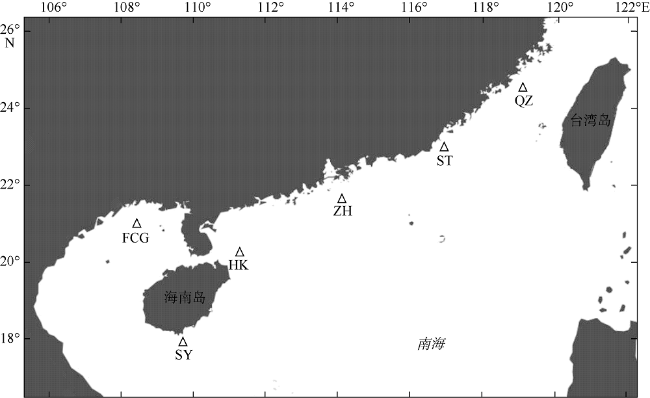
图1 花斑蛇鲻样本的采集点示意图该图基于“百度地图个性在线编辑器”中审图号为GS(2019)5218号的标准地图制作 Fig. 1 Map of sampling sites for S. undosquamis |
表1 花斑蛇鲻样本信息及D-loop区序列遗传多样性参数Tab. 1 Specimen information of S. undosquamis and genetic diversity parameters based on D-loop sequences |
| 地理群体(缩写) | 采样点位置 | 样本量/个 | 单倍型数量/个 | 单倍型多样性(h±SD) | 核苷酸多样性(π±SD) |
|---|---|---|---|---|---|
| 防城港(FCG) | 108°30'E, 21°00'N | 21 | 19 | 0.9905 ± 0.0178 | 0.0110 ± 0.0058 |
| 三亚(SY) | 109°46'E, 17°58'N | 22 | 21 | 0.9957 ± 0.0153 | 0.0148 ± 0.0077 |
| 海口(HK) | 111°18'E, 20°18'N | 22 | 21 | 0.9957 ± 0.0153 | 0.0135 ± 0.0071 |
| 珠海(ZH) | 114°05'E, 21°41'N | 21 | 20 | 0.9952 ± 0.0165 | 0.0142 ± 0.0074 |
| 汕头(ST) | 116°55'E, 23°00'N | 22 | 19 | 0.9827 ± 0.0208 | 0.0129 ± 0.0068 |
| 泉州(QZ) | 119°02'E, 24°36'N | 21 | 21 | 1.0000 ± 0.0147 | 0.0128 ± 0.0068 |
| 总计 | / | 129 | 101 | 0.9873 ± 0.0048 | 0.0132 ± 0.0067 |
注: SD为标准差(Standard Deviation) |
图2 花斑蛇鲻D-loop区序列单倍型邻接树的拓扑结构及在地理群体中的分布各分支标记大于70%的自展值(百分数, 图中未标示百分号); Saurida wanieso代表外群鳄蛇鲻; 带框的单倍型为共享单倍型; 各颜色代表不同地理群体来源的单倍型, 数字代表单倍型频率 Fig. 2 Topology of NJ tree for S. undosquamis D-loop sequence haplotypes, and distribution of haplotypes among geographic populations. Bootstrap values of >70% are shown at nodes (percentage). Saurida wanieso is used as outgroup. The shared haplotypes are framed. The colors represent the corresponding geographic populations, and the numbers represent the haplotype frequencies |
表2 花斑蛇鲻6个地理群体D-loop区序列遗传变异的分子方差分析Tab. 2 Analysis of molecular variance for six populations of S. undosquamis based on D-loop sequences |
| 变异来源 | 自由度(无量纲) | 变异百分比/% | 分化指数(Fst) | P值 | |
|---|---|---|---|---|---|
| 群体间 | 5 | 0.13 | / | / | |
| 群体内 | 123 | 99.87 | / | / | |
| 所有样本 | 128 | / | 0.0013 | 0.3992 | |
注:“/”表示无此项 |
表3 花斑蛇鲻两两地理群体间D-loop区序列的遗传分化指数(对角线下方)及显著性水平(对角线上方)Tab. 3 Pairwise FST (below diagonal) and P values (above diagonal) among geographic populations of S. undosquamis based on D-loop sequences |
| FCG | SY | HK | ZH | ST | QZ | |
|---|---|---|---|---|---|---|
| FCG | 0.0773 | 0.0707 | 0.0202 | 0.1060 | 0.1605 | |
| SY | 0.0269 | 0.9300 | 0.6829 | 0.8408 | 0.7405 | |
| HK | -0.0267 | -0.0211 | 0.7952 | 0.9263 | 0.8341 | |
| ZH | 0.0478 | -0.0126 | -0.0155 | 0.9421 | 0.2658 | |
| ST | -0.0506 | -0.0176 | -0.0204 | -0.0238 | 0.3424 | |
| QZ | 0.0164 | -0.0155 | -0.0178 | 0.0071 | 0.0017 |
表4 花斑蛇鲻两两地理群体间随机交配假设检验的显著性水平Tab. 4 P values of exact test of sample differentiation of S. undosquamis based on D-loop haplotype frequencies |
| FCG | SY | HK | ZH | ST | |
|---|---|---|---|---|---|
| SY | 0.0146 | ||||
| HK | 0.0351 | 0.7368 | |||
| ZH | 0.2855 | 0.2921 | 0.1818 | ||
| ST | 0.3906 | 0.7042 | 0.8730 | 0.6746 | |
| QZ | 0.5270 | 0.3033 | 0.4864 | 0.3926 | 0.9453 |
| [1] |
蔡研聪, 徐姗楠, 陈作志, 等, 2018. 南海北部近海渔业资源群落结构及其多样性现状[J]. 南方水产科学, 14(2):10-18.
|
| [2] |
陈再超, 刘继兴, 1982. 南海经济鱼类[M]. 广州: 广东科学与技术出版社: 184-188(in Chinese).
|
| [3] |
陈作志, 邱永松, 徐姗楠, 等, 2012. 北部湾花斑蛇鲻生物学特征的演化[J]. 中国水产科学, 19(2):321-328.
|
| [4] |
郜星晨, 章群, 薛丹, 等, 2016. 基于线粒体控制区部分序列的南海大斑石鲈遗传多样性分析[J]. 海洋科学, 40(7):41-45.
|
| [5] |
黄小林, 李文俊, 林黑着, 等, 2018. 基于线粒体DNA D-loop序列的黄斑篮子鱼群体遗传多样性分析[J]. 热带海洋学报, 37(4):45-51.
|
| [6] |
黄梓荣, 2002. 休渔对南海北部多齿蛇鲻资源的影响[J]. 湛江海洋大学学报, 22(6):26-31.
|
| [7] |
江艳娥, 许友伟, 范江涛, 等, 2019. 南海北部陆架水域多齿蛇鲻与花斑蛇鲻的年龄与生长[J]. 中国水产科学, 26(1):82-90.
|
| [8] |
梁述章, 宋炜, 马春艳, 等, 2019. 基于线粒体控制区的中国近海棘头梅童鱼群体遗传结构研究[J]. 海洋渔业, 41(2):138-148.
|
| [9] |
吕金磊, 章群, 杨喜书, 等, 2017. 基于线粒体控制区的中国南海海域卵形鲳鲹遗传多样性[J]. 海洋渔业, 39(3):241-248.
|
| [10] |
舒黎明, 邱永松, 2004a. 南海北部花斑蛇鲻生长死亡参数估计及开捕规格[J]. 湛江海洋大学学报, 24(3):29-35.
|
| [11] |
舒黎明, 邱永松, 2004b. 南海北部多齿蛇鲻生物学分析[J]. 中国水产科学, 11(2):154-158.
|
| [12] |
苏纪兰, 袁业立, 2005. 中国近海水文[M]. 北京: 海洋出版社: 1-367(in Chinese).
|
| [13] |
孙冬芳, 董丽娜, 李永振, 等, 2010. 南海北部海域多齿蛇鲻的种群分析[J]. 水产学报, 34(9):1387-1394.
|
| [14] |
孙典荣, 林昭进, 2004. 北部湾主要经济鱼类资源变动分析及保护对策探讨[J]. 热带海洋学报, 23(2):62-68.
|
| [15] |
熊丹, 李敏, 陈作志, 等, 2015. 南海短尾大眼鲷的种群遗传结构分析[J]. 南方水产科学, 11(2):27-34.
|
| [16] |
许友伟, 陈作志, 范江涛, 等, 2015. 南沙西南陆架海域底拖网渔获物组成及生物多样性[J]. 南方水产科学, 11(5):76-81.
|
| [17] |
杨炳忠, 杨吝, 谭永光, 等, 2017. 南海区2种桁杆虾拖网渔获组成调查与分析[J]. 南方水产科学, 13(6):115-122.
|
| [18] |
杨喜书, 章群, 薛丹, 等, 2018. 中国近海细鳞鯻线粒体控制区的遗传多样性[J]. 生态学报, 38(5):1852-1859.
|
| [19] |
张俊, 陈国宝, 陈作志, 等, 2015. 南沙南部陆架海域渔业资源声学评估[J]. 南方水产科学, 11(5):1-10.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}