
基于DNA条形码研究中国枪乌贼和鸢乌贼的食物种类组成*
|
刘梦娜(1992—), 女, 河南省焦作市人, 在读硕士, 研究方向为海洋生物学。E-mail: |
Copy editor: 姚衍桃
收稿日期: 2019-10-29
要求修回日期: 2020-01-19
网络出版日期: 2020-07-27
基金资助
国家科技基础资源调查专项(2017FY201405)
中国水产科学研究院基本科研业务费资助(2019HY-JC0203)
广东省渔业生态环境重点实验室开放基金(FEEL-2019-9)
版权
Study on food contents of Uroteuthis chinensis and Sthenoteuthis oualaniensis based on COI sequence
Received date: 2019-10-29
Request revised date: 2020-01-19
Online published: 2020-07-27
Supported by
Foundation item: The Science and Technology Project of Basic Resources of China(2017FY201405)
Basal Research Fund of Chinese Academy of Fishery Sciences(2019HY-JC0203)
Open Foundation of Key Laboratory of Fishery Ecology Environment of Guangdong Province(FEEL-2019-9)
Copyright
中国枪乌贼(Uroteuthis chinensis)和鸢乌贼(Sthenoteuthis oualaniensis)作为中国南海头足类的关键种, 对海洋生态系统的物质流动与能量循环具有重要影响, 其摄食生态的研究将对海洋食物网的构建具有重要意义。由于传统胃含物分析法难以准确鉴定糜状饵料生物的组成, 本研究利用DNA条码技术, 针对中国枪乌贼和鸢乌贼不可辨认的食物糜提取组织DNA, 选用线粒体基因细胞色素C氧化酶亚基I(Mitochondrial cytochrome oxidase subunit I, COI)作为分子标记, 获得的序列在Genbank中进行比对分析, 并使用GMYC(Generalized Mixed Yule Coalescent)模型进行物种界定和构建系统进化关系。结果成功鉴定出中国枪乌贼饵料物种有13种, 鸢乌贼饵料物种有8种, 共20种(其中1种为共有饵料)。比较发现, 汕头-台湾浅滩渔场的中国枪乌贼主要摄食鱼类、甲壳类和头足类, 而南海中部海域鸢乌贼则主要摄食鱼类和头足类。两物种均存在同类相食现象, 但鸢乌贼表现更明显。
刘梦娜 , 徐磊 , 王雪辉 , 刘玉 , 王淼娣 , 邱永松 , 朱江峰 , 何映霖 , 贝伟烈 , 杜飞雁 . 基于DNA条形码研究中国枪乌贼和鸢乌贼的食物种类组成*[J]. 热带海洋学报, 2020 , 39(4) : 61 -69 . DOI: 10.11978/2019105
Uroteuthis chinensis and Sthenoteuthis oualaniensis are key species of the cephalopods in the South China Sea, which have important effects on the material flow and energy circulation in the marine ecosystem. The study of feeding ecology of these two cephalopods will be great significant to the construction of marine food web. Because of the sharp tooth decay, the stomach contents are all scorpion-like, and it is difficult to accurately identify them by traditional morphological methods. And the effectiveness of molecular techniques in feeding analysis of these two species has not been fully validated. In this study, we identified the species (or genus) in the stomach contents of U. chinensis and S. oualaniensis based on the mitochondrial COI gene, and used the Generalized Mixed Yule Coalescent (GMYC) model for species definition and phylogenetic tree building. The results showed that of a total of 41 stomach contents sequences (28 from U. chinensis and 13 from S. oualaniensis), 30 (73%) were identified as species, and the rest were determined as genus (all are crustaceans). The diets of U. chinensis in Shantou-Taiwan Shoal fishing ground included fish, cephalopods, and crustaceans; and the diets of S. oualaniensis in the Central South China Sea were fish and cephalopods. Cannibalism occurred in both species, but was more pronounced in S. oualaniensis.
表1 鸢乌贼和中国枪乌贼采样站位表Tab. 1 Sanpling sites of S. oualaniensis and U. chinensis in the South China Sea |
| 站位 | 东经 | 北纬 | |
|---|---|---|---|
| 鸢乌贼 | S3 | 111°37'50" | 14°58'04" |
| S9 | 110°35'06" | 14°02'10" | |
| S16 | 117°28'28" | 13°00'00" | |
| S18 | 110°30'35" | 11°02'09" | |
| S19 | 111°28'51" | 10°59'30" | |
| S20 | 113°51'30" | 11°02'17" | |
| S21 | 114°32'36" | 11°06'13" | |
| S26 | 110°28'22" | 9°30'53" | |
| 中国枪乌贼 | SQ | 118°29'21" | 23°05'46" |
表2 PCR引物信息Tab. 2 Information of PCR primers |
| 引物名称 | 序列(5′-3′) | 参考文献 |
|---|---|---|
| HCO2198(下游) | TAAACTTCAGGGTGACCAAAAAATCA | Folmer et al, 1994 |
| dgHCO2198(下游) | TAAACTTCAGGGTGACCAAARAAYCA | Meyer, 2003 |
| jgHCO2198(下游) | TAIACYTCIGGRTGICCRAARAAYCA | Geller et al, 2013 |
| mlCOIintF(上游) | GGWACWGGWTGAACWGTWTAYCCYCC | Leray et al, 2013 |
表3 2018年7月汕头-台湾浅滩渔场海域中国枪乌贼食物组成Tab. 3 Taxonomic distribution of diets of U. chinensis in Shantou-Taiwan shoal fishing ground in July, 2018 |
| 最佳匹配物种 | 相似度 | 序列号 | 样品号 |
|---|---|---|---|
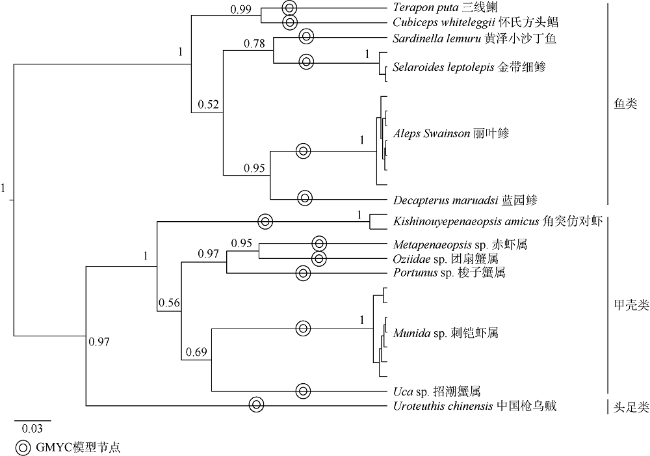
| 克氏副葉鰺 Alepes Swainson | 99.90% | KY371115.1 | N0709、N0711、N0752、N0759、N0767、N0771、N0772 |
| 蓝圆鲹 Decapterus maruadsi | 100.00% | MK264594.1 | N0718 |
| 金带细鲹 Selaroides leptolepis | 99.64% | JX261038.1 | N0740、N0742、N0779 |
| 三线鯻 Terapon puta | 100.00% | KY372209.1 | N0722 |
| 怀氏方头鲳 Cubiceps whiteleggii | 100.00% | KY371367.1 | N0743 |
| 黄泽小沙丁鱼 Sardinella lemuru | 100.00% | MK264430.1 | N0758 |
| 角突仿对Kishinouyepenaeopsis amicus | 98.71% | KR349255.1 | N0720、N0723 |
| 赤虾属 Metapenaeopsis sp. | 87.05% | KR150460.1 | N0753 |
| 刺铠虾属 Munida sp. | 86.58% | AY350938.1 | N0749、N0754、N0757、N0761、N0768、N0769、N0775 |
| 招潮蟹属 Uca sp. | 84.40% | AB813678.1 | N0735 |
| 团扇蟹属 Ozius sp. | 87.45% | HM638053.1 | N0770 |
| 梭子蟹属Portunus sp. | 88.67% | JX398096.1 | N0777 |
| 中国枪乌贼 Uroteuthis chinensis | 100.00% | MG192385.1 | N0736 |
注: 重复出现的饵料物种(属)的相似度为平均取值。 |
图2 基于COI基因序列构建的中国枪乌贼饵料生物贝叶斯系统进化树Fig. 2 Bayes phylogenetic tree of diet species (genus) of U. chinensis base on mitochondria COI gene sequences |
表4 2019 年7 月南海中部海域鸢乌贼食物组成Tab. 4 Taxonomic distribution of diets of S. oualaniensis in the South China Sea in July, 2019 |
| 最佳匹配物种 | 相似度 | 序列号 | 样品号 |
|---|---|---|---|
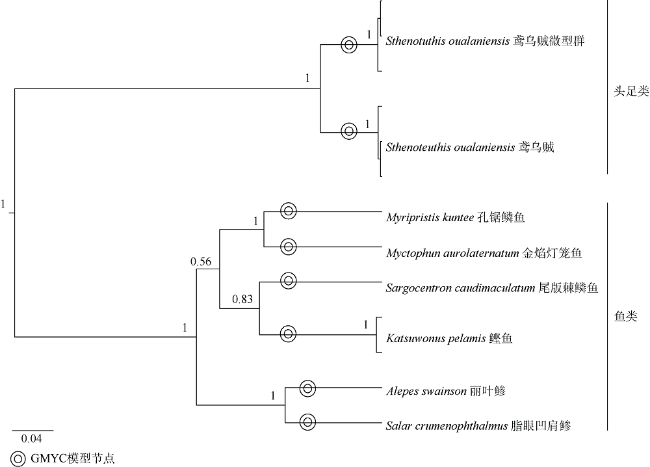
| 孔锯鳞鱼 Myripristis kuntee | 99.53% | MK658063.1 | S20-41-2 |
| 尾斑棘鳞鱼 Sargocentron caudimaculatum | 100.00% | MF409503.1 | S19-44-4 |
| 克氏副葉鰺 Alepes Swainson | 100.00% | MK264421.1 | S3-23-3 |
| 脂眼凹肩鲹 Selar crumenophthalmus | 100.00% | MH638727.1 | S26-14-2 |
| 鲣鱼 Katsuwonus pelamis | 99.78% | MH638766.1 | S19-44-3、S16-24-3 |
| 金焰灯笼魚 Myctophum aurolaternatum | 100.00% | KJ555427.1 | S20-28-3 |
| 鸢乌贼微型群 Sthenoteuthis oualaniensis | 100.00% | MF411130.1 | S18-1、S18-14、S19-31-2 |
| 鸢乌贼 Sthenoteuthis oualaniensis | 100.00% | MF411105.1 | S21-45-1、S3-25、S9-22 |
注: 重复出现的饵料物种(属)的相似度为平均取值。由于目前尚未通过分子生物学方法对鸢乌贼进行物种区分命名, 表中仅采用形态学上普遍使用的名称对鸢乌贼的中文名称进行了区分, 而拉丁文名称则未进行区分。 |
| [1] |
陈大纲, 张美昭, 2015. 中国海洋鱼类[M]. 青岛: 中国海洋大学出版社: 280-1883(in Chinese).
|
| [2] |
陈新军, 2004. 渔业资源与渔场学[M]. 北京: 海洋出版社: 51-52(in Chinese).
|
| [3] |
陈新军, 刘必林, 王尧耕, 2009. 世界头足类[M]. 北京: 海洋出版社: 312-313(in Chinese).
|
| [4] |
戴泉水, 卢振彬, 洪明进, 等, 2004. 台湾海峡南部海域游泳生物区系组成和渔业资源现状[J]. 中国水产科学, 11(4):360-366.
|
| [5] |
董正之, 1988. 中国动物志, 软体动物门, 头足纲[M]. 北京: 科学出版社: 88-95(in Chinese).
|
| [6] |
范江涛, 冯雪, 邱永松, 等, 2013. 南海鸢乌贼生物学研究进展[J]. 广东农业科学, 40(23):122-128.
|
| [7] |
龚金科, 1981. 闽南-台湾浅滩渔场台湾枪乌贼的洄游分布和生物学特性的探讨[J]. 福建水产科技, ( 2):15-26 (in Chinese).
|
| [8] |
龚玉艳, 詹凤娉, 杨玉滔, 等, 2016. 南海鸢乌贼摄食习性的初步研究[J]. 南方水产科学, 12(4):80-87.
|
| [9] |
龚玉艳, 孔啸兰, 杨玉滔, 等, 2018. 南海鸢乌贼微型群体的摄食习性研究[J]. 海洋渔, 40(4):394-403.
|
| [10] |
黄美珍, 2004. 台湾海峡及邻近海域4种头足类的食性和营养级研究[J]. 台湾海峡, 23(3):331-340.
|
| [11] |
李斌, 陈国宝, 郭禹, 等, 2016. 南海中部海域渔业资源时空分布和资源量的水声学评估[J]. 南方水产科学, 12(4):28-37.
|
| [12] |
唐启升, 1999. 海洋食物网与高营养层次营养动力学研究策略[J]. 海洋水产研究, 20(2):1-6.
|
| [13] |
颜云榕, 冯波, 卢伙胜, 等, 2012. 南沙群岛北部海域鸢乌贼(Sthenoteuthis oualaniensis)夏季渔业生物学研究[J]. 海洋与湖沼, 43(6):1177-1186.
|
| [14] |
张宇美, 颜云榕, 卢伙胜, 等, 2013. 西沙群岛海域鸢乌贼摄食与繁殖生物学初步研究[J]. 广东海洋大学学报, 33(3):56-64.
|
| [15] |
张壮丽, 叶孙忠, 洪明进, 等, 2008. 闽南—台湾浅滩渔场中国枪乌贼生物学特性研究[J]. 福建水产, 3(1):1-5.
|
| [16] |
郑玉水, 杨甘霖, 曾建忠, 等. 台湾海峡枪乌贼资源探捕调查报告[J]. 福建水产. 1988: 8-16.
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}