
南海珊瑚礁海域黑缘尾九棘鲈生物学特征初步研究
|
洪小帆(1996—), 男, 硕士研究生, 研究方向为海洋渔业生态。email: |
Copy editor: 姚衍桃
收稿日期: 2020-07-06
修回日期: 2020-08-21
网络出版日期: 2020-08-21
基金资助
国家自然科学基金项目(31902374)
国家重点研发计划资助(2018YFC1406502)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0605)
农业农村部财政专项(NFZX2021)
版权
Biological characteristics of Cephalopholis spiloparaea of reef waters in the South China Sea
Copy editor: YAO Yantao
Received date: 2020-07-06
Revised date: 2020-08-21
Online published: 2020-08-21
Supported by
National Natural Science Foundation of China(31902374)
National Key Research and Development Program of China(2018YFC1406502)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0605)
Financial Fund of the Ministry of Agriculture and Rural Affairs(NFZX2021)
Copyright
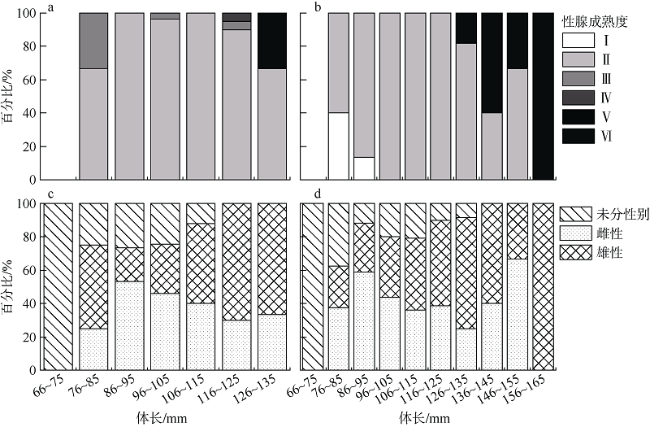
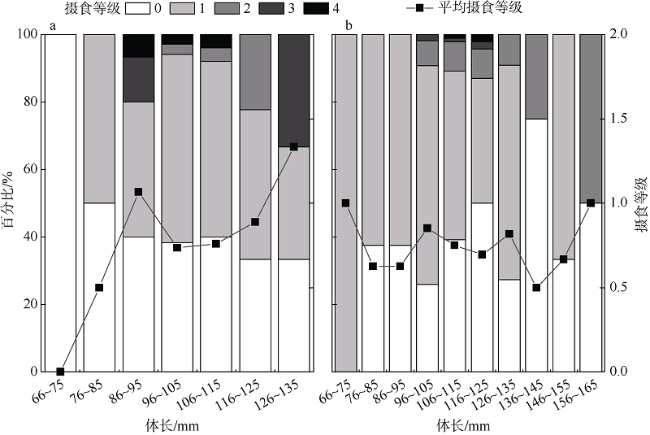
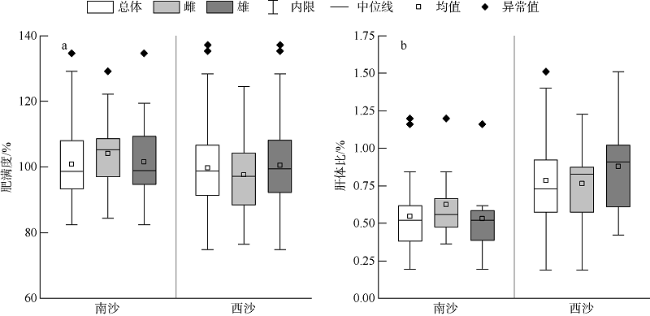
根据2018—2019年在南沙群岛和西沙群岛珊瑚礁海域采集的礁栖鱼类黑缘尾九棘鲈(Cephalopholis spiloparaea)样本, 对其群体结构、性腺成熟度、摄食强度、肥满度、年龄与生长等生物学特征进行了初步研究, 通过对比分析了南海这两个珊瑚礁海域中黑缘尾九棘鲈生物学特征的地理差异。结果表明: 在南沙海域和西沙海域中黑缘尾九棘鲈的平均体长分别为105.5±10.5mm和110.4±13.1mm, 两海域间的体长和体质量分布均具有极显著的差异(P<0.01); 南沙海域与西沙海域样本体长和体质量关系的异速生长因子b值分别为2.881与2.797; 耳石样本分析结果显示年龄范围为0.58~1.44a, 两个海域间并无显著性差异(P>0.05); 由von Bertalanffy生长方程计算所得的生长参数显示, 南沙海域的黑缘尾九棘鲈比西沙海域的体型相对较小、生长速度相对较快; 两海域黑缘尾九棘鲈的性腺成熟度均以Ⅱ期为主, 且西沙海域样本性腺成熟比例高于南沙海域; 摄食等级以1级为主, 两个海域间的摄食等级分布无显著性差异(P>0.05); 与西沙群岛海域相比, 南沙群岛海域的黑缘尾九棘鲈群体表现出肝体比较低的特征。
洪小帆 , 陈作志 , 江艳娥 , 张俊 , 王欢欢 , 李媛洁 , 李纲 . 南海珊瑚礁海域黑缘尾九棘鲈生物学特征初步研究[J]. 热带海洋学报, 2021 , 40(4) : 50 -62 . DOI: 10.11978/2020069
Based on the coral reef fishes of Cephalopholis spiloparaea (Serranidae: Cephalopholis), which were collected from the coral reef waters of the Nansha Islands and Xisha Islands in the South China Sea in September 2018 and May-June 2019, we studied the biological characteristics of C. spiloparaea, including population structure, gonad maturity composition, feeding intensity, fatness index, age, and growth. We compared the geographic variation of the biological characteristics between the two coral reef waters in the South China Sea. The results are as follows. 1) The ranges of the dominant body length were 96~125 mm and 96~115 mm, respectively, in the Nansha reef waters and Xisha reef waters. The distributions of body length and body weight both had significant differences between the two waters (P<0.01). 2) The allometric growth factors (b values) of the body length and body weight relationship from the Nansha reef waters and Xisha reef waters were 2.881 and 2.797, respectively. 3) The result of analysis of otolith samples (May-June 2019) suggests that the age range is 0.58~1.44 a. There were no significant differences in the distribution of age between the two reef waters (P>0.05). 4) The C. spiloparaea in the Nansha reef waters was relatively smaller in size and grew faster than that in the Xisha reef waters. 5) In addition, the gonadal maturation ratio of the Xisha reef waters was higher than that of the Nansha reef waters. 6) The feeding stages of C. spiloparaea from the two reef waters were dominated in Stage 1. Compared with the population of C. spiloparaea in the Xisha reef waters, these results demonstrate that the population in the Nansha reef waters had the characteristics of miniaturization, faster growth rate, and lower hepatopancreas index.
Key words: Cephalopholis spiloparaea; coral reef fish; biology; South China Sea
表1 南海南沙群岛和西沙群岛海域黑缘尾九棘鲈样本采集信息Tab. 1 Sampling information of C. spiloparaea specimens from Nansha Islands and Xisha Islands waters in the South China Sea |
| 年份 | 地区 | 岛礁 | 样本数目 | 采样方法 |
|---|---|---|---|---|
| 2018 | 南沙群岛 | 美济礁、永暑礁、渚碧礁 | 47 | 手钓 |
| 2018 | 西沙群岛 | 琛航岛 | 110 | 手钓 |
| 2019 | 南沙群岛 | 永暑礁、渚碧礁 | 61 | 手钓、地笼网 |
| 2019 | 西沙群岛 | 琛航岛、七连屿 | 144 | 手钓、地笼网 |
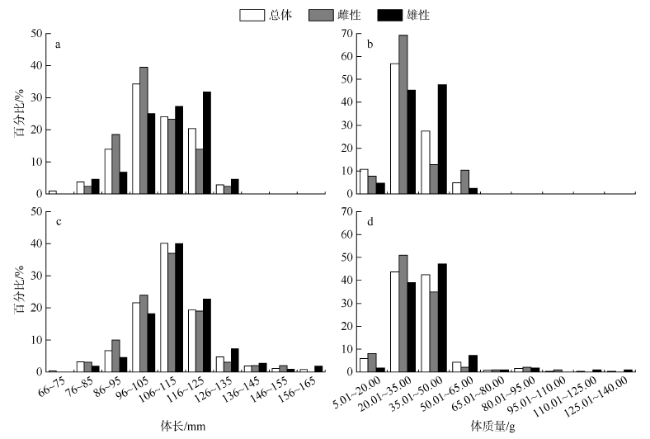
图1 黑缘尾九棘鲈的体长、体质量分布a、b: 南沙海域; c、d: 西沙海域 Fig. 1 Distributions of body length and body weight of C. spiloparaea |
表2 2018—2019年南沙和西沙海域黑缘尾九棘鲈的体长、体质量信息Tab. 2 Information of body length and body weight of C. spiloparaea in Nansha and Xisha waters in 2018 and 2019 |
| 海域 | 群体 | 体长范围/mm | 平均体长/mm | 优势体长组/mm | 体质量范围/g | 平均体质量/g | 优势体质量范围/g |
|---|---|---|---|---|---|---|---|
| 南沙 | 总体 | 70~134 | 105.5±10.5 | 96~125 | 9.30~58.89 | 32.03±10.49 | 20.01~50.00 |
| 雌性 | 84~134 | 104.7±10.9 | 96~115 | 16.84~58.89 | 32.10±10.88 | 20.01~35.00 | |
| 雄性 | 83~128 | 108.9±10.3 | 96~125 | 15.21~54.32 | 35.22±9.28 | 20.01~50.00 | |
| 西沙 | 总体 | 68~162 | 110.4±13.1 | 96~115 | 8.46~138.36 | 36.11±14.67 | 20.01~50.00 |
| 雌性 | 80~152 | 109.3±12.7 | 96~115 | 13.08~101.87 | 34.35±13.62 | 20.01~50.00 | |
| 雄性 | 78~162 | 113.1±13.4 | 106~125 | 12.6~138.36 | 38.98±16.74 | 20.01~50.00 |
表3 黑缘尾九棘鲈的体长-体质量关系参数Tab. 3 Estimated parameters of body length vs body weight of C. spiloparaea in Nansha and Xisha Islands waters |
| 群体 | 南沙海域 | 西沙海域 | 总体 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| a(×10-5) | b | R2 | a(×10-5) | b | R2 | a(×10-5) | b | R2 | |
| 雌性 | 5.436 | 2.851 | 0.9068 | 5.625 | 2.829 | 0.8892 | 6.892 | 2.789 | 0.8872 |
| 雄性 | 6.677 | 2.801 | 0.8732 | 9.031 | 2.735 | 0.882 | 8.673 | 2.744 | 0.8818 |
| 总群体 | 4.566 | 2.881 | 0.9108 | 6.685 | 2.797 | 0.8945 | 6.277 | 2.811 | 0.9009 |
注: R2为决定系数(Coefficient of Determination) |
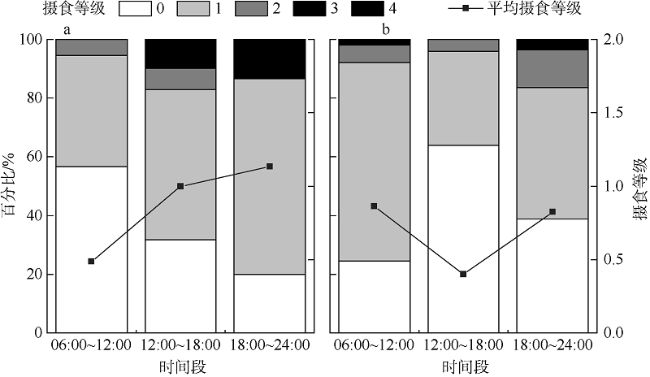
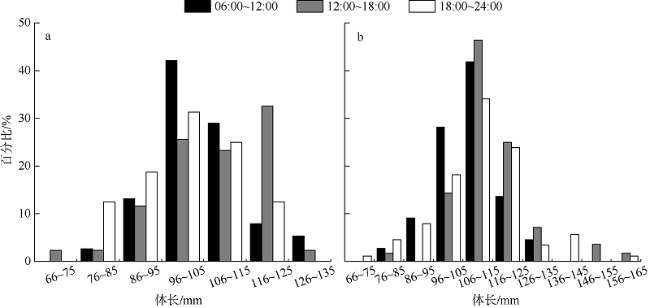
图4 黑缘尾九棘鲈不同时间段的摄食等级百分比a. 南沙海域; b. 西沙海域 Fig. 4 Percentage of different feeding stages of C. spiloparaea at different times: a) samples from Nansha waters; b) samples from Xisha waters |
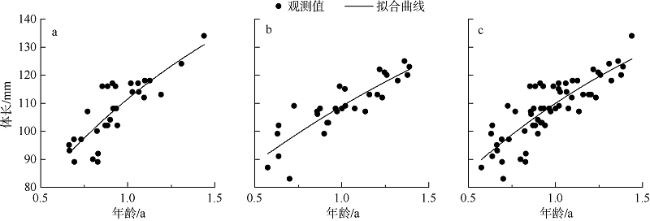
图7 黑缘尾九棘鲈的体长-年龄关系a. 永暑礁(2019年5月); b. 琛航岛(2019年6月); c. 总体 Fig. 7 Relationship between body length and age of C. spiloparaea |
表4 南沙和西沙海域黑缘尾九棘鲈的生长参数Tab. 4 Growth parameters of C. spiloparaea in reef areas of the South China Sea |
| 拟合参数 | L∞/mm | t0/a | k/a-1 | RSS | R2 |
|---|---|---|---|---|---|
| 南沙海域 | 185.24 | -0.33 | 0.69 | 1128 | 0.68 |
| 西沙海域 | 195.44 | -0.91 | 0.42 | 769 | 0.73 |
| 总体 | 174.78 | -0.56 | 0.64 | 2074 | 0.69 |
注: RSS为残差平方和(Residual Sum of Squares); R2为决定系数(Coefficient of Determination) |
| [1] |
曹希全, 李艳慧, 魏杰, 等, 2019. 宽口裂腹鱼体长-体质量关系和肥满度[J]. 西北农业学报, 28(8):1380-1386.
|
| [2] |
陈国宝, 李永振, 2005. 南海主要珊瑚礁鮨科鱼类的组成与分布[J]. 南方水产, 1(3):18-25.
|
| [3] |
陈新军, 2014. 渔业资源与渔场学[M]. 2版. 北京: 海洋出版社: 1-460(in Chinese).
|
| [4] |
戴强, 戴建洪, 李成, 等, 2006. 关于肥满度指数的讨论[J]. 应用与环境生物学报, 12(5):715-718.
|
| [5] |
单秀娟, 胡芷君, 邵长伟, 等, 2020. 捕捞诱导鱼类生物学特征进化研究进展[J]. 渔业科学进展, 41(3):165-175.
|
| [6] |
费鸿年, 张诗全, 1990. 水产资源学[M]. 北京: 中国科学技术出版社.
|
| [7] |
傅秀梅, 邵长伦, 王长云, 等, 2009. 中国珊瑚礁资源状况及其药用研究调查Ⅱ. 资源衰退状况、保护与管理[J]. 中国海洋大学学报, 39(4):685-690.
|
| [8] |
高永利, 黄晖, 练健生, 等, 2014. 西沙群岛礁栖鱼类物种多样性及其食性特征[J]. 生物多样性, 22(5):618-623.
|
| [9] |
郜星晨, 姜伟, 张琪, 等, 2019. 长江宜昌段鳙的繁殖生物学初步研究[J]. 水生态学杂志, 40(6):92-98.
|
| [10] |
贾丹丹, 2018. 基于遥感的南海珊瑚礁区海表温度时空变化及生态影响的研究[D]. 南宁: 广西大学.
|
| [11] |
江艳娥, 陈作志, 林昭进, 等, 2019. 南海海域鸢乌贼中型群与微型群渔业生物学比较[J]. 水产学报, 43(2):454-466.
|
| [12] |
李建华, 张鑫浩, 金岳, 等, 2018. 基于耳石和角质颚微结构的中国枪乌贼年龄与生长比较[J]. 海洋渔业, 40(5):513-521.
|
| [13] |
李娇, 公丕海, 常青, 等, 2020. 岩礁鱼类行为生态学研究进展[J]. 渔业科学进展, 41(6):192-199.
|
| [14] |
李淑, 余克服, 陈天然, 等, 2011. 珊瑚共生虫黄藻密度结合卫星遥感分析2007年南沙群岛珊瑚热白化[J]. 科学通报, 56(10):756-764.
|
| [15] |
李永振, 贾晓平, 陈国宝, 2007. 南海珊瑚礁鱼类资源[M]. 北京: 海洋出版社 (in Chinese).
|
| [16] |
李忠炉, 金显仕, 单秀娟, 等, 2011. 小黄鱼体长-体质量关系和肥满度的年际变化[J]. 中国水产科学, 18(3):602-610.
|
| [17] |
李忠义, 杨尧尧, 应一平, 等, 2018. 南海北部陆坡金鼻眶灯鱼秋季摄食节律[J]. 中国水产科学, 25(2):395-402.
|
| [18] |
刘必林, 陈新军, 钟俊生, 2009. 采用耳石研究印度洋西北海域鸢乌贼的年龄、生长和种群结构[J]. 大连海洋大学学报, 24(3):206-212.
|
| [19] |
刘必林, 陈新军, 李建华, 2016. 东太平洋茎柔鱼的耳石微结构[J]. 水产学报, 40(8):1211-1217.
|
| [20] |
刘飞, 韦慧, 胡隐昌, 等, 2019. 流溪河入侵鱼类豹纹脂身鲶的年龄与生长特征研究[J]. 淡水渔业, 49(5):22-30.
|
| [21] |
龙丽娟, 杨芳芳, 韦章良, 2019. 珊瑚礁生态系统修复研究进展[J]. 热带海洋学报, 38(6):1-8.
|
| [22] |
庞旭, 2014. 基于温度变化的鱼类生理生态(热耐受、代谢及游泳)响应研究[D]. 重庆: 重庆大学.
|
| [23] |
区又君, 柳琪, 刘泽伟, 2006. 3种笛鲷的含肉率、肥满度、比肝重和肌肉营养成分的分析[J]. 大连水产学院学报, 21(3):287-289.
|
| [24] |
邵广昭, 陈正平, 陈静怡, 等, 2011. 南海东沙岛及太平岛鱼类种类组成和动物地理学特点[J]. 生物多样性, 19(6):737-763.
|
| [25] |
王耕, 王希哲, 2019. 渔业扩张与珊瑚礁白化之间的关联性分析——以南海珊瑚礁为例[J]. 辽宁师范大学学报(自然科学版), 42(4):518-524.
|
| [26] |
王继隆, 刘伟, 王维坤, 等, 2020. 绥芬河大麻哈鱼个体生物学研究[J]. 水生生物学报, 44(1):162-169.
|
| [27] |
王婷, 刘利平, 陈桃英, 等, 2015. 鳗鲡(Anguilla japonica)性腺发育和饥饿胁迫下生物学指标及体内蛋白质与氨基酸含量变化[J]. 海洋与湖沼, 46(6):1373-1379.
|
| [28] |
韦众, 鲍传和, 1999. 不同水体河鲀丰满度、性腺和肝脏指数的比较[J]. 安徽农业大学学报, 26(4):427-431.
|
| [29] |
谢小军, 邓利, 张波, 1998. 饥饿对鱼类生理生态学影响的研究进展[J]. 水生生物学报, 22(2):181-188.
|
| [30] |
许莉莉, 薛莹, 徐宾铎, 等, 2018. 海州湾大泷六线鱼摄食生态研究[J]. 中国水产科学, 25(3):608-620.
|
| [31] |
杨骏, 何兴恒, 孙治宇, 2020. 中华花鳅的生长和繁殖生物学的研究[J]. 水产科学, 39(2):209-217.
|
| [32] |
殷名称, 1995. 鱼类生态学[M]. 北京: 中国农业出版社 (in Chinese).
|
| [33] |
詹秉义, 2005. 渔业资源评估[M]. 北京: 中国农业出版社 (in Chinese).
|
| [34] |
张国政, 李显森, 朱建成, 等, 2010. 黄海中南部小黄鱼当年幼鱼生长特征的研究[J]. 渔业科学进展, 31(3):15-22.
|
| [35] |
张辉, 姜亚洲, 袁兴伟, 等, 2020. 象山港黄姑鱼增殖群体早期生态适应特征[J]. 中国水产科学, 27(1):118-125.
|
| [36] |
张俊, 陈作志, 陈国宝, 2015. 南沙海域永暑礁金带齿颌鲷生物学特征初步研究[J]. 南方水产科学, 11(5):108-116.
|
| [37] |
张俊, 陈作志, 董俊德, 等, 2020. 近20年南海四带笛鲷(Lutjanus kasmira)种群特征变化[J]. 海洋与湖沼, 51(1):114-124.
|
| [38] |
张魁, 陈作志, 邱永松, 2016. 北部湾二长棘犁齿鲷生长、死亡和性成熟参数的年际变化[J]. 南方水产科学, 12(6):9-16.
|
| [39] |
赵焕庭, 王丽荣, 袁家义, 2016. 南海诸岛珊瑚礁可持续发展[J]. 热带地理, 36(1):55-65.
|
| [40] |
朱玲, 温海深, 毛玉泽, 2002. 野生鲇性腺发育及其与肝重指数关系的研究[J]. 水利渔业, 22(5):26-27.
|
| [41] |
朱庆环, 田永军, 张弛, 等, 2020. 基于耳石微结构的黄、渤海蓝点马鲛幼鱼日龄与生长[J]. 海洋学报, 42(2):87-95.
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}