
大型海藻龙须菜凋落物分解对水质的影响
|
戴晓娟(1995—), 女, 广东省陆丰市人, 硕士研究生, 从事海洋生态与大型海藻修复研究。email: |
Copy editor: 姚衍桃
收稿日期: 2020-09-02
要求修回日期: 2020-11-25
网络出版日期: 2020-11-19
基金资助
国家自然科学基金项目(41977268)
版权
Effects of the decomposition of Gracilaria lemaneiformis on seawater quality
Received date: 2020-09-02
Request revised date: 2020-11-25
Online published: 2020-11-19
Supported by
National Natural Science Foundation of China(41977268)
Copyright
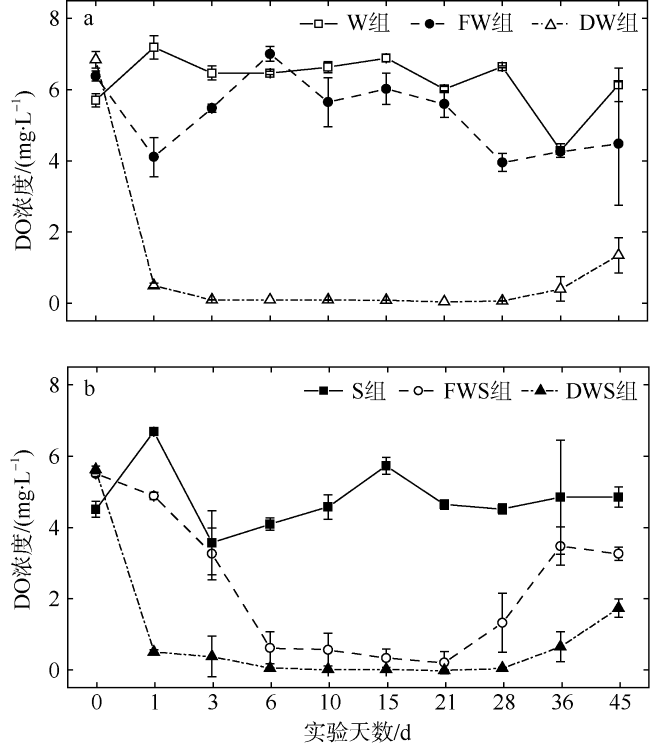
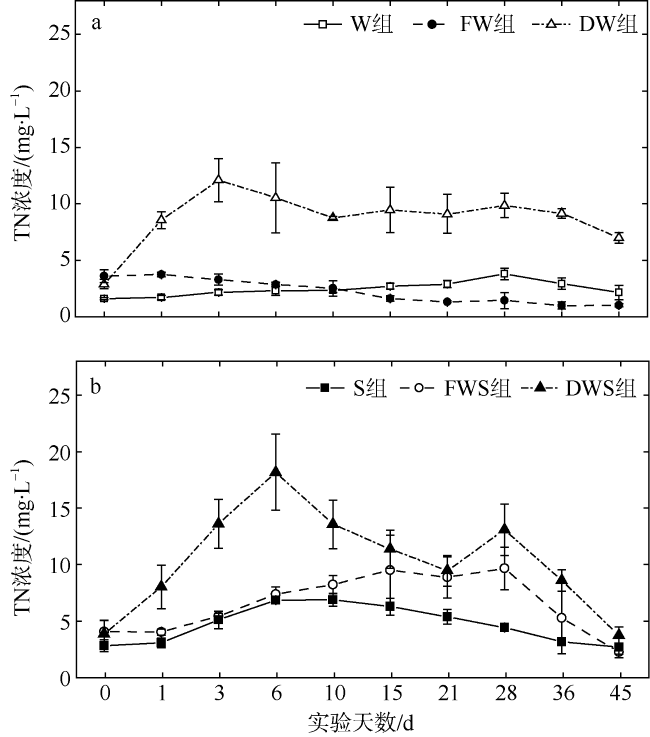
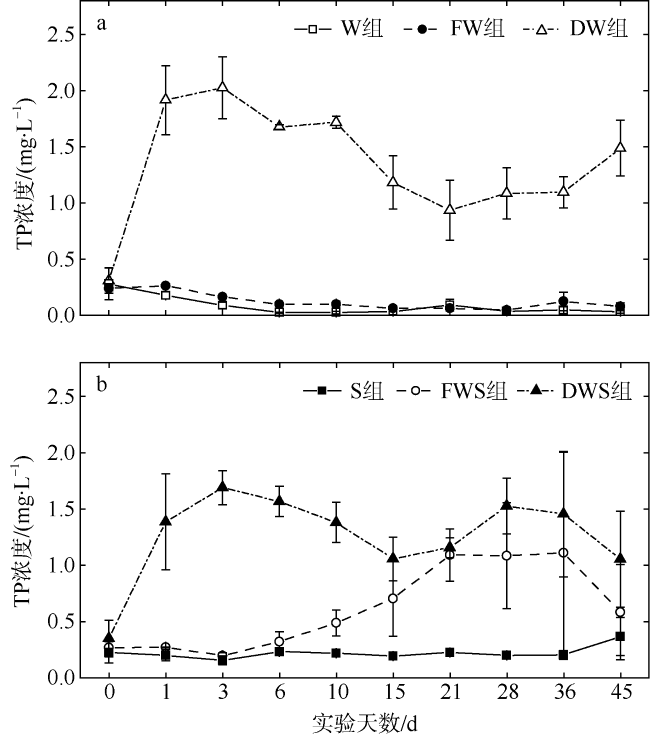
大型海藻龙须菜(Gracilaria lemaneiformis)规模栽培具有重要的经济和环境效益, 但藻体的凋落分解会对栽培区和邻近海域水环境造成一定影响。为探讨龙须菜凋落对水环境的影响, 本文通过45d的室内受控实验, 评估了龙须菜凋落分解过程中水体溶解氧和氮、磷的含量变化。结果发现, 干龙须菜实验组在实验期内水体溶解氧浓度显著降低(较对照组降低了82.81%); 水体氮、磷浓度显著提高, 总氮、总磷浓度较对照组分别上升了161.78%和759.93%。鲜龙须菜+海水+沉积物组在实验前中期(第0~21天)水体溶解氧浓度持续降低(较对照组降低了53.92%), 但在21d后又逐渐恢复至对照组水平; 其水体氮、磷浓度在实验中末期亦显著提高, 分解过程总氮、总磷浓度分别较对照组上升了36.65%和177.80%, 水体氮、磷变化曲线较干龙须菜组平缓且迟滞。鲜龙须菜凋落分解过程中的营养盐释放率低于干龙须菜, 沉积物对鲜龙须菜的分解及氮、磷和碳释放有促进作用, 但对干龙须菜的分解及氮、磷和碳释放有一定程度的减缓作用。龙须菜失重率、分解速率及营养盐释放率均呈现如下规律: 干龙须菜+海水组>干龙须菜+海水+沉积物组>鲜龙须菜+海水+沉积物组>鲜龙须菜+海水组。依据上述结果, 建议在龙须菜规模栽培和收获过程中应及时打捞脱落或衰老藻体, 尤其对已收获的大型海藻应妥善处理, 避免大型海藻腐烂而导致水体污染。
戴晓娟 , 胡韧 , 罗洪添 , 王庆 , 胡晓娟 , 白敏冬 , 杨宇峰 . 大型海藻龙须菜凋落物分解对水质的影响[J]. 热带海洋学报, 2021 , 40(1) : 91 -98 . DOI: 10.11978/2020099
Large-scale cultivation of seaweed Gracilaria lemaneiformis has important economic and environmental benefits. However, the litter decomposition of the seaweed may affect water quality in the cultivation area and its adjacent waters. To investigate the effects of the decomposition of G. lemaneiformis on water quality, a decomposition experiment was carried out for 45 days. The results showed that the concentration of dissolved oxygen (DO) reduced 82.81% in the dried seaweed group compared with the control group, and the concentrations of total nitrogen and phosphorus increased 161.78% and 759.93%, respectively. While DO concentrations in the treatment of fresh seaweed+seawater+sediment (FWS) continuously decreased from 5.56mg·L-1 to 0.26mg·L-1 from Day 0 to 21, and then gradually recovered to the control level at the end of the experiment. Significantly increase of nitrogen (36.65%) and phosphorus (177.80%) concentrations in water bodies were also observed in FWS treatment in the mid-late period of the experiment. Meanwhile, the change curves of nitrogen and phosphorus in fresh seaweed treatment were slower and gentler than those of the dried seaweed treatment. The nutrients release rates of fresh seaweed were lower than those of dried seaweed during the litter decomposition process. Sediment promoted the decomposition of fresh seaweed and the release rates of carbon, nitrogen and phosphorus, but had opposite effects on dried seaweed. The weight loss rate and the decomposition rate and the nutrients release rates of G. lemaneiformis followed a descending order of dried seaweed+seawater treatment (DW), dried seaweed+seawater+sediment treatment (DWS), fresh seaweed+seawater+sediment treatment (FWS), and fresh seaweed+seawater treatment (FW). To avoid secondary pollution to the sea water, we suggest that shedding or senescent algae be removed in time during the large-scale cultivation period, and the harvested seaweed should be properly deposed.
Key words: macroalgae; Gracilaria lemaneiformis decomposition; litter; nutrient; sediments
图1 龙须菜凋落物分解实验设置W组表示海水组; S组表示海水+沉积物组; DW组表示干龙须菜+海水组; FW组表示鲜龙须菜+海水组; DWS组表示干龙须菜+海水+沉积物组; FWS组表示鲜龙须菜+海水+沉积物组 Fig. 1 Experimental design of G. lemaneiformis litter decomposition W represents seawater, S represents sediment, DW represents dried seaweed+seawater, FW represents fresh seaweed+seawater, DWS represents dried seaweed+seawater+sediment, and FWS represents fresh seaweed+seawater+sediment |
表1 龙须菜的失重率、分解速率和营养盐释放率Tab. 1 The weight loss rate, decomposition rate and release rate of nutrients in G. lemaneiformis |
| 组别 | 失重率/% | 分解速率/d-1 | TN释放率/% | TP释放率/% | TC释放率/% |
|---|---|---|---|---|---|
| 鲜龙须菜+海水组(FW组) | 12.42±3.55b | 0.003±0.001b | -1.13±7.38b | 15.95±4.90b | 6.96±6.49b |
| 鲜龙须菜+海水+沉积物组(FWS组) | 62.50±13.39ab | 0.02±0.01ab | 53.18±12.86ab | 58.38±14.35ab | 56.84±15.39ab |
| 干龙须菜+海水组(DW组) | 89.58±3.03a | 0.05 ± 0.01a | 73.96±5.61a | 86.0±4.70a | 86.20±3.32a |
| 干龙须菜+海水+沉积物组(DWS组) | 81.04±12.67a | 0.04±0.02a | 57.06±22.34ab | 81.70±8.16a | 80.80±10.11a |
注: 数值为平均值±标准差; 同列数据右上角不同小写字母表示不同实验组之间有显著差异(P<0.05) |
表2 龙须菜TN、TP单位累积释放量Tab. 2 Unit cumulative release of TN and TP from G. lemaneiformis |
| 组别 | TN/(mg·g-1) | TP/(mg·g-1) |
|---|---|---|
| 鲜龙须菜+海水组(FW组) | -1.81 | 0.33 |
| 干龙须菜+海水组(DW组) | 52.35 | 10.50 |
| 鲜龙须菜+海水+沉积物组(FWS组) | 14.84 | 3.20 |
| 干龙须菜+海水+沉积物组(DWS组) | 47.36 | 8.68 |
| [1] |
曹勋, 韩睿明, 章婷曦, 等, 2015. 冬季水生植物分解过程及其对水质的影响研究[J]. 农业环境科学学报, 34(2):361-369.
|
| [2] |
陈洪森, 叶春, 李春华, 等, 2020. 入湖河口区水生植物群落衰亡分解释放营养盐过程模拟研究[J]. 环境工程技术学报, 10(2):220-228.
|
| [3] |
杜立刚, 2013. 三峡库区消落带植被淹水碳氮磷释放及消落带氮磷交换通量研究[D]. 重庆: 重庆大学.
|
| [4] |
刘湘庆, 王宗灵, 辛明, 等, 2016. 浒苔衰亡过程中营养盐的释放过程及规律[J]. 海洋环境科学, 35(6):801-805, 813.
|
| [5] |
刘之威, 罗洪添, 武宇辉, 等, 2019. 汕头南澳龙须菜规模栽培对水质和浮游植物的影响[J]. 中国水产科学, 26(1):99-107.
|
| [6] |
孟祥森, 邵雪琳, 高丽, 等, 2016. 绿潮硬毛藻衰亡分解过程中营养盐的释放规律[J]. 海洋环境科学, 35(4):495-500.
|
| [7] |
农业农村部渔业渔政管理局, 2019. 中国渔业年鉴[M]. 北京: 中国农业出版社.
Fisheries and Fisheries Administration Bureau of the Ministry of Agriculture and Rural Areas, 2019. China fisheries yearbook[M]. Beijing: China Agricultural Press (in Chinese).
|
| [8] |
童雄, 罗沛, 刘锋, 等, 2019. 绿狐尾藻分解及其氮磷释放特征[J]. 环境科学, 40(7):3118-3125.
|
| [9] |
王博, 叶春, 杨劭, 等, 2009. 腐解黑藻生物量对高硝态氮水体氮素的影响[J]. 环境科学研究, 22(10):1198-1203.
|
| [10] |
王立志, 王国祥, 2013. 衰亡期沉水植物对水和沉积物磷迁移的影响[J]. 生态学报, 33(17):5426-5437.
|
| [11] |
王立志, 宋红丽, 董彬, 等, 2020. 黑藻与金鱼藻自然衰亡过程中营养盐释放规律研究[J]. 环境科学研究, 33(1):138-146.
|
| [12] |
王云祥, 李正, 秦传新, 等, 2016. 不同季节江蓠脱落物对大型海藻场上覆水的影响[J]. 南方水产科学, 12(2):13-20.
|
| [13] |
叶春, 王博, 李春华, 等, 2014. 沉水植物黑藻腐解过程中营养盐释放过程[J]. 中国环境科学, 34(10):2653-2659.
|
| [14] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.4-2007 海洋调查规范第4部分: 海水化学要素调查[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration , 2008. GB/T 12763.4-2007 Specifications for oceanographic survey-Part 4: Survey of chemical parameters in sea water[S]. Beijing: Standards Press of China (in Chinese).
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}