
海水升温对非虫黄藻共生型珊瑚Cladopsammia sp.的生理影响*
|
张喆(1997—), 女, 山西省晋中市人, 硕士研究生, 从事珊瑚生物学和珊瑚礁生态学研究。email: |
Copy editor: 殷波
收稿日期: 2021-10-19
修回日期: 2021-12-21
网络出版日期: 2021-12-22
基金资助
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0402)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0404)
Effect of ocean warming on the physiology of azooxanthellate coral–Cladopsammia sp.*
Copy editor: YIN Bo
Received date: 2021-10-19
Revised date: 2021-12-21
Online published: 2021-12-22
Supported by
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0402)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0404)
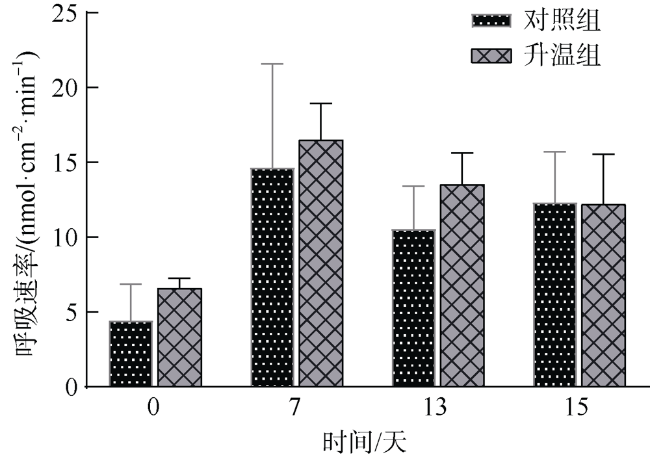
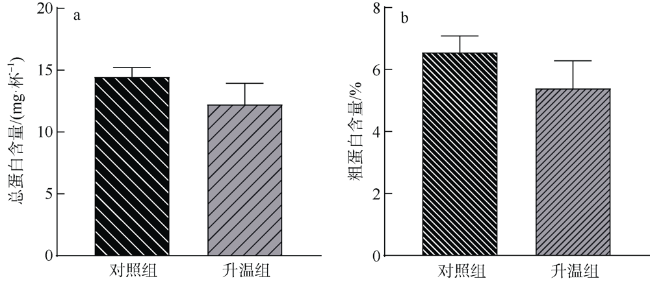
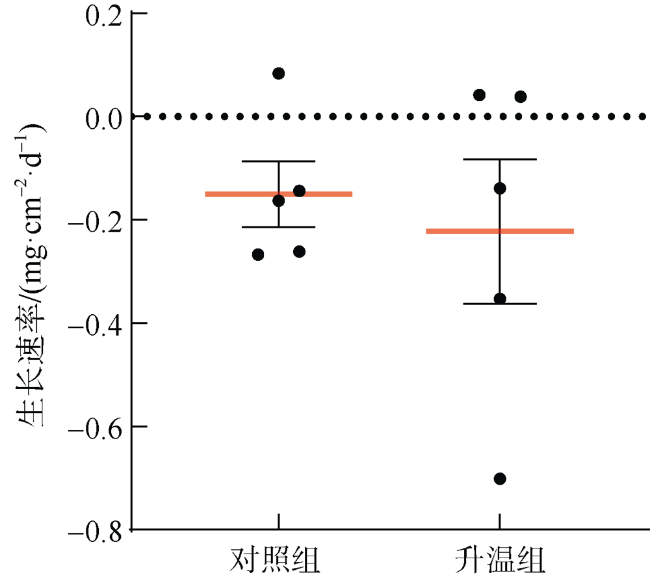
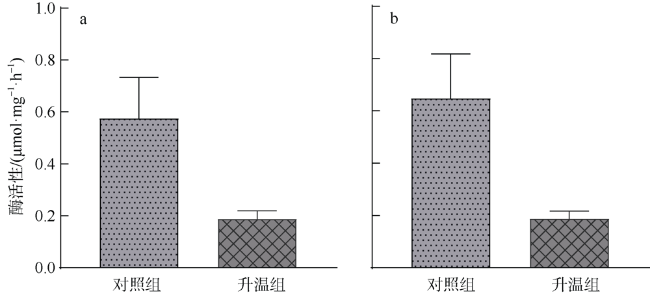
文章以非虫黄藻共生型珊瑚Cladopsammia sp.为研究对象, 将对照组的海水设置为26℃恒温, 试验组从26℃升温至33℃, 以此探究海水升温对Cladopsammia sp.代谢和钙化生理的影响。结果显示: 在高温胁迫下, 虽然Cladopsammia sp.钙化相关酶(Ca2+-ATP酶和Mg2+-ATP酶)的活性出现了负响应, 但是珊瑚的呼吸速率、生长速率、组织中总蛋白和粗脂肪都没有显著改变(p>0.05)。比较历史文献结果和本研究结果表明: Cladopsammia sp.可能由于体内没有虫黄藻共生, 表现出了比大部分虫黄藻共生珊瑚更高的温度耐受能力。
关键词: 海水升温; 非虫黄藻共生型珊瑚Cladopsammia sp.; 代谢; 钙化
张喆 , 俞晓磊 , 梁宇娴 , 何茜 , 黄晖 , 袁翔城 , 周伟华 . 海水升温对非虫黄藻共生型珊瑚Cladopsammia sp.的生理影响*[J]. 热带海洋学报, 2022 , 41(5) : 141 -149 . DOI: 10.11978/2021140
To study the effects of ocean warming on the metabolism and calcification physiology of azooxanthellate coral - Cladopsammia sp., we set the seawater in controlling group at 26 ℃, and the temperature of experimental group was increased from 26 ℃ to 33 ℃. The results indicated that although the activity of calcification-related enzymes (Ca2+-ATPase and Mg2+-ATPase) of Cladopsammia sp. showed a negative response, the respiration rate, growth rate, total protein and crude lipid in tissues of this coral were not significantly altered under thermal stress (p > 0.05). The comparison between the data of previous studies and this study shows that Cladopsammia sp. exhibited higher temperature tolerance than most zooxanthellate corals, probably because it does not have Symbiodinium. At present, the research on the thermal stress of azooxanthellate corals is very limited. It is necessary to strengthen the research on the mechanism of tolerance of these corals, to help us predict more accurately the impact of future environmental changes on coral reef ecosystem in China.
表1 不同时间的实验温度设置Tab. 1 Experimental temperature at different times |
| 时间 | 第1天(D0) | 第7天(D7) | 第13天(D13) | 第15天(D15) |
|---|---|---|---|---|
| 对照组温度/℃ | 26±0.5 | 26±0.5 | 26±0.5 | 26±0.5 |
| 试验组温度/℃ | 26±0.3 | 29±0.3 | 32±0.3 | 33±0.3 |
表2 文献记录的海洋钙化物种在不同试验处理后的Ca2+-ATP酶活性和Mg2+-ATP酶活性Tab. 2 The activity of Ca2+-ATPase and Mg2+-ATPase of marine calcifying species recorded in literatures after different experimental treatments |
| 海洋钙化物种 | 是否虫黄藻共生 | 试验处理 | Ca2+-ATP酶活性/(μmol·mg-1·h-1) | Mg2+-ATP酶活性/(μmol·mg-1·h-1) | 参考文献 |
|---|---|---|---|---|---|
| Mussismilia harttii (珊瑚) | 是 | 铜富集试验培养12天(6.7μg·L-1) | 2.52 ± 0.45 | 3.85 ± 0.29 | de Barros Marangoni等(2017) |
| Tubastrea aurea (珊瑚) | 否 | 自然条件 | 0.23 | 0.52 | Liao等(2021) |
| Cladopsammia sp. (珊瑚) | 否 | 自然条件 | 0.88±0.16 | 0.92±0.16 | 本研究 |
| Marginopora vertebralis (底栖有孔虫) | 否 | 自然条件 | 0.63 | 1.11 | Prazeres等(2015) |
| [1] |
国家海洋信息中心, 2022. 中国气候变化海洋蓝皮书(2021)[M]. 北京: 科学出版社.
NATIONAL MARINE DATA INFORMATION CENTER, 2022. Blue book on marine climate change in China 2021[M]. Beijing: Science Press. (in Chinese)
|
| [2] |
郭亚娟, 周伟华, 袁翔城, 等, 2018. 两种造礁石珊瑚对海水酸化和溶解有机碳加富的响应[J]. 热带海洋学报, 37(1): 57-63.
|
| [3] |
焦尚, 霸婉玉, 2017. 动物海产品中粗脂肪测定方法的研究[J]. 现代盐化工, 44(2): 28-29, 31.
|
| [4] |
梁宇娴, 俞晓磊, 郭亚娟, 等, 2020. 3种传统方法对不同珊瑚表面积测量的适用性及其校准方法—以3D扫描技术为基准[J]. 热带海洋学报, 39(1): 85-93.
|
| [5] |
亚力士, 2018. 三亚湾近岸海域及其珊瑚生长区重金属污染现状与评价[D]. 南宁: 广西大学: 1-73.
|
| [6] |
俞晓磊, 江雷, 罗勇, 等, 2019. 异养营养对丛生盔形珊瑚代谢及共生藻光合生理的影响[J]. 海洋科学, 43(12): 81-88.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE, 2019. Special report on the ocean and cryosphere in a changing climate[R]. Geneva: IPCC.
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}