
不同光强下长茎葡萄蕨藻(Caulerpa lentillifera)直立枝和匍匐枝的光生理特征及其对升温的响应
|
史晓寒(1997—), 女, 河南省周口市, 硕士研究生, 从事海洋藻类生理生态及应用研究。email: |
Copy editor: 殷波
收稿日期: 2021-09-27
修回日期: 2021-12-22
网络出版日期: 2021-12-28
基金资助
广州市科技计划(202206010159)
南方海洋科学与工程广东省实验室(珠海)资助项目(SML2021SP203)
国家自然科学基金(41890853)
Photophysiological characteristics of the branch and stolon of macroalga Caulerpa lentillifera (Caulerpaceae, Caulerpa) under different growth light conditions, and their responses to temperature rise
Copy editor: YIN Bo
Received date: 2021-09-27
Revised date: 2021-12-22
Online published: 2021-12-28
Supported by
Science and Technology Planning Project of Guangzhou(202206010159)
Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai)(SML2021SP203)
National Natural Science Foundation(41890853)
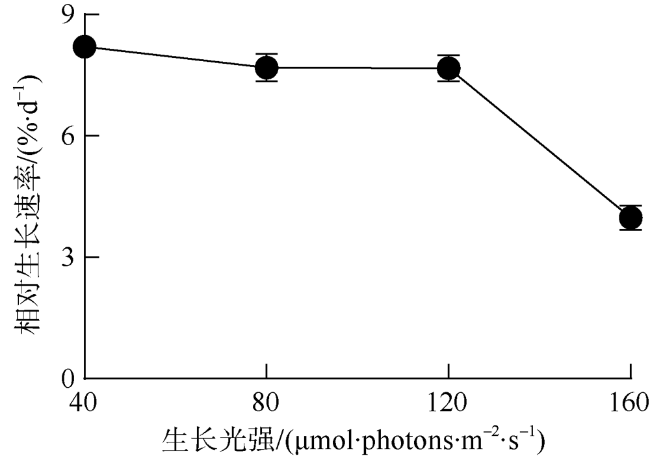
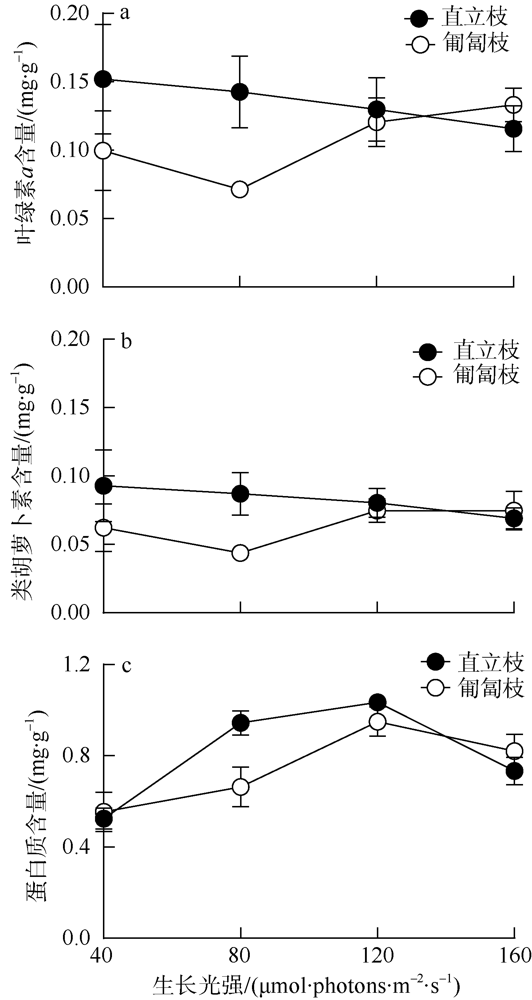
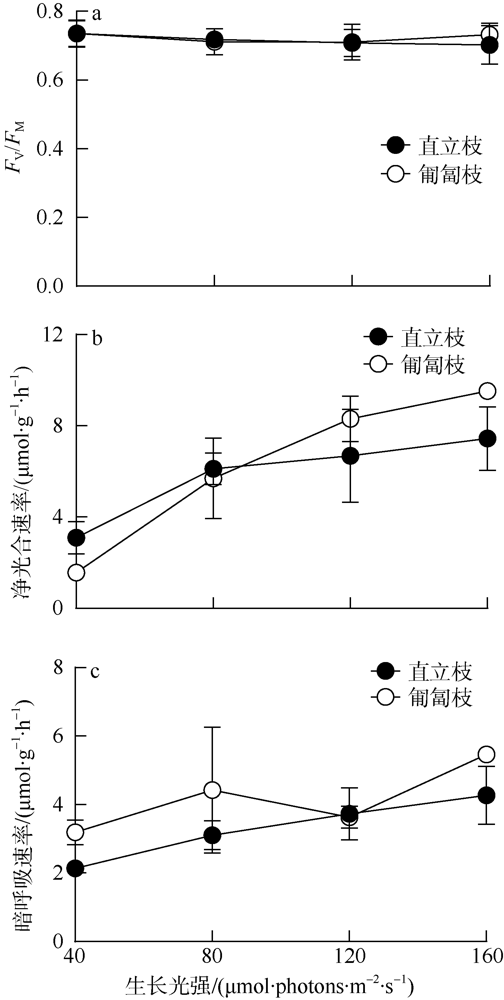
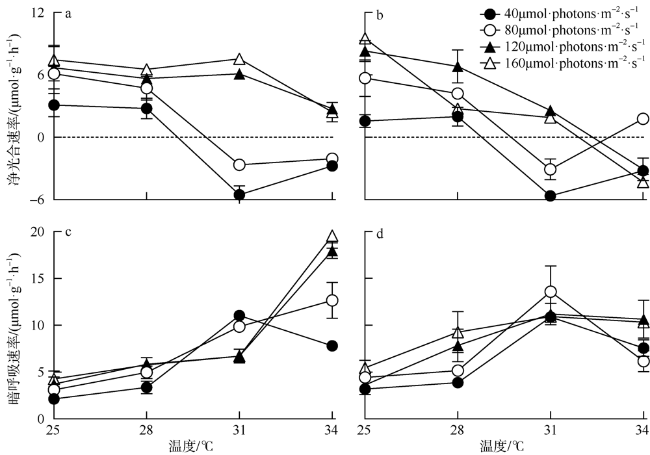
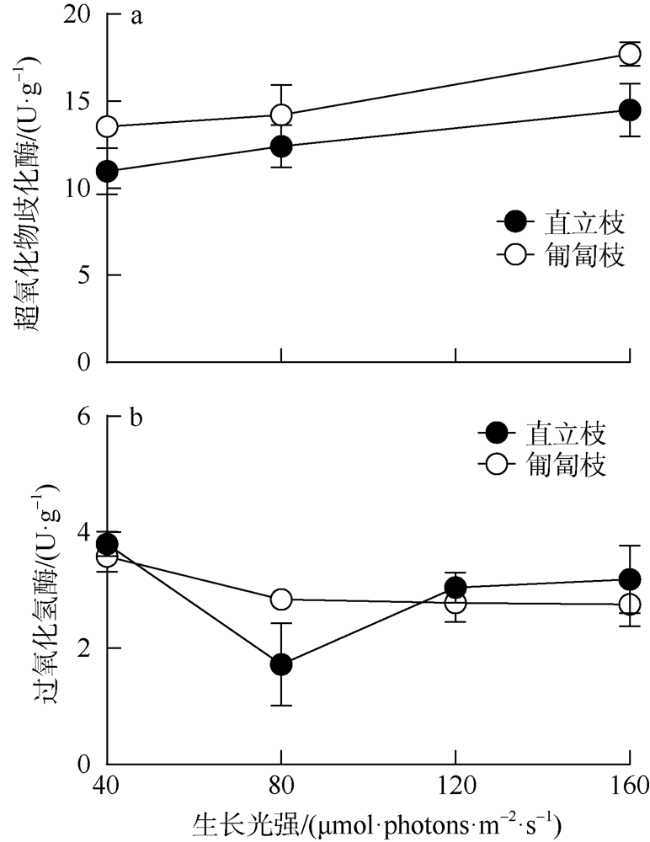
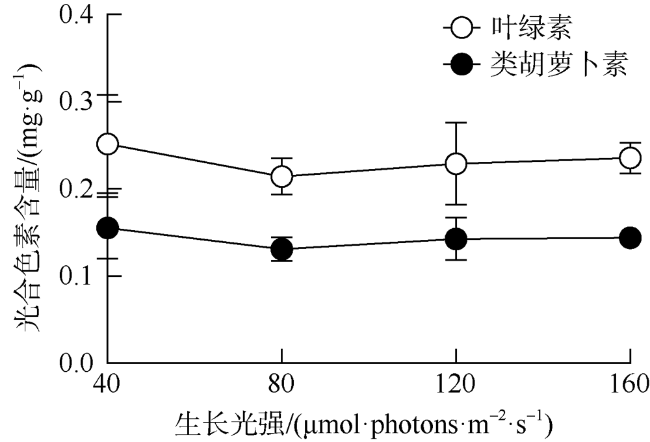
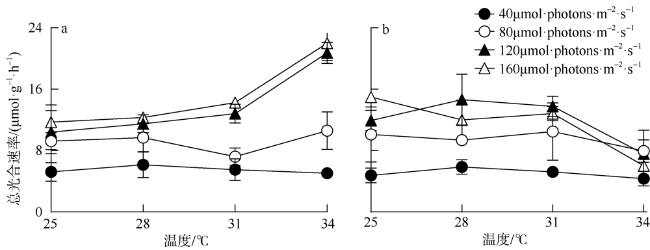
海洋绿藻长茎葡萄蕨藻(Caulerpa lentillifera, 又名海葡萄)因具有较高经济和生态价值而备受关注, 光照和温度变化均会改变长茎葡萄蕨藻生理代谢, 最终影响其经济价值和生态功能。文章比较研究不同生长光强下(40、80、120和160 µmol·photons·m-2·s-1)长茎葡萄蕨藻不同部位, 即直立枝和匍匐枝的生理和生化特征, 以及其对升温(+3℃、+6℃和+9℃)的响应。结果显示, 光强由40升至120µmol·photons·m-2·s-1时对长茎葡萄蕨藻相对生长率(RGR)的影响不显著, 但是光强升至160µmol·photons·m-2·s-1时可使RGR降低49%。弱光下(40µmol·photons·m-2·s-1)直立枝的叶绿素(Chl a)和类胡萝卜素(Car)含量为匍匐枝的1.52和1.49倍; 直立枝的Chl a和Car含量随生长光强升高而降低, 匍匐枝随光强升高而升高, 二者蛋白含量则均随光强升高而先升高后降低。弱光下直立枝的净光合放氧速率(Pn)和呼吸速率(Rd)分别为匍匐枝的2倍和70%, 但是二者的最大光化学效率(FV/FM)差异不显著。光强升高提高直立枝和匍匐枝的Pn和Rd, 但对二者FV/FM的影响不显著。同时, 弱光下直立枝的超氧化物歧化酶(SOD)活性比匍匐枝低20%, 二者过氧化氢酶(CAT)活性差异不显著; 光强升高提高直立枝和匍匐枝的SOD活性, 降低CAT活性。研究还发现, 直立枝和匍匐枝的Pn随温度升高而降低, 但前者的降低程度即光合速率随升温的变化率随光强升高而降低, 后者的则随光强升高而升高, 可见温度升高在弱光下对长茎葡萄蕨藻直立枝的负面影响更大, 在强光下则对匍匐枝的负面影响更大。
史晓寒 , 邹定辉 , 何泉 , 李刚 . 不同光强下长茎葡萄蕨藻(Caulerpa lentillifera)直立枝和匍匐枝的光生理特征及其对升温的响应[J]. 热带海洋学报, 2022 , 41(5) : 150 -160 . DOI: 10.11978/2021132
Marine green macroalga Caulerpa lentillifera (Caulerpaceae, Caulerpa) has attracted significant attention because of its high economic and ecological values. Changes in light and temperature influence the algal physiological metabolism and consequently affect its economic value and ecological function. In this study, biochemical compositions and physiological characteristics of the branch and stolon of C. lentillifera were comparably explored under light intensities of 40, 80, 120 and 160 µmol·photons·m-2·s-1, as well as their photosynthetic responses to temperature rise (+3, +6 and +9 ℃) under each light level. The results showed that varying light intensities from 40 to 120 µmol·photons·m-2·s-1 had no significant effect on the relative growth rate [i.e., RGR, (7.85 ± 0.26) %·d-1], but the growth light of 160 µmol·photons·m-2·s-1 reduced the RGR by 49 %. Under 40 µmol·photons·m-2·s-1 light intensity, chlorophyll a (Chl a) and carotenoids (Car) concentrations of branch were (0.15 ± 0.04) and (0.093 ± 0.020) mg·g-1, being about 1.52- and 1.49-fold of stolon; both Chl a and Car of branch increased with increasing light intensities, but that of stolon decreased. Protein contents of both branch and stolon increased from low to medium growth lights, then decreased to high light, with the maximum values of (1.03 ± 0.00) and (0.95 ± 0.06) mg·g-1 under 120 µmol·photons·m-2·s-1, respectively. Under 40 µmol·photons·m-2·s-1 light intensity, the photosynthetic oxygen evolution rate (Pn), dark respiration rate (Rd) and photosynthetic efficiency (FV/FM) of branch were (3.10 ± 0.71), (2.14 ± 0.09) µmol·g-1·h-1 and (0.74 ± 0.04), wherein the Pn and Rd were 2-fold and 70 % of stolon, but no significant difference in the FV/FM between them. Both Pn and Rd increased with rising growth lights, but the FV/FM did not change. Superoxide dismutase (SOD) activity in branch [(11.0 ± 1.32) U·g-1] was about 20 % lower than that in stolon under 40 µmol·photons·m-2·s-1 growth light, while the catalase (CAT) activity [(3.80 ± 0.21) U·g-1] was similar. The SOD activity increased with increasing growth lights in both branch and stolon, but the CAT activity decreased. Particularly, our results demonstrated that the temperature rise reduced the Pn of both branch and stolon, with the reduction degree (i.e., the slope of Pn versus temperature) decreased with growth lights in branch, but increased in stolon, indicating the temperature rise maybe more harmful to branch in low light and to stolon in high light conditions.
图3 不同生长光强下长茎葡萄蕨藻直立枝和匍匐枝FV/FM (a)、光合速率(b)和暗呼吸速率(c)变化曲线Fig. 3 Maximal photochemical quantum yield (FV/FM) of Photosystem II (a), photosynthetic oxygen evolution rate (b) and dark respiration rate (c) in the branch and stolon of Caulerpa lentillifera under various growth light intensities |
| [1] |
程丽巍, 邹定辉, 郑青松, 等, 2010. 光照和温度对氮饥饿及饱和营养条件下石莼(Ulva lactuca)的硝态氮吸收动力学影响[J]. 生态学杂志, 29(5): 939-944.
|
| [2] |
丁柳丽, 邹定辉, 刘露, 等, 2015. 气候变化对海藻龙须菜生长与光合作用耐热特性的影响[J]. 生态学报, 35(10): 3267-3277.
|
| [3] |
董潇潇, 靳红磊, 王宏斌, 2016. 植物光系统高光适应机制研究进展[J]. 植物生理学报, 52(11): 1725-1732.
|
| [4] |
高坤山, 2014. 藻类固碳—理论、进展与方法[M]. 北京: 科学出版社: 1-491.
|
| [5] |
高秀秀, 李亚鹤, 段维军, 等, 2015. 光强和二氧化碳浓度变化对浒苔幼苗生长及生理的影响[J]. 海洋学报, 37(10): 80-87.
|
| [6] |
郭辉, 2014. 海葡萄(Caulerpa lentillifera)切断组织再生培养及发育条件研究[D]. 青岛: 中国科学院研究生院(海洋研究所).
|
| [7] |
黄丹, 刘东超, 王晓梁, 等, 2019. 不同温度下长茎葡萄蕨藻无机碳利用[J]. 广东海洋大学学报, 39(3): 61-69.
|
| [8] |
姜芳燕, 宋文明, 杨宁, 等, 2014. 长茎葡萄蕨藻的人工养殖技术研究[J]. 热带农业科学, 34(8): 99-103.
|
| [9] |
兰志刚, 李新仲, 肖钢, 等, 2016. 海上浮式核电站温排水对海洋生态环境的影响[J]. 海洋科学, 40(6): 84-88.
|
| [10] |
李红, 党晨阳, 张金荣, 2018. 三种马尾藻不同部位挥发性成分的比较分析[J]. 食品工业科技, 39(24): 281-288, 293.
|
| [11] |
柳波, 孙彬, 马家海, 2003. 经济海藻资源的开发利用[J]. 渔业现代化, (3): 35-36.
|
| [12] |
聂修和, 聂宜茂, 聂俊华, 等, 1992. 光合有效辐射测量原理及其单位换算[J]. 山东农业大学学报(自然科学版), 23(3): 247-253, 258.
|
| [13] |
沈国英, 黄凌风, 郭丰, 等, 2010. 海洋生态学[M]. 北京: 科学出版社: 1-360.
|
| [14] |
施建宏, 2008. 台湾蕨藻之调查与养殖研究[D]. 中国台湾: 国立中山大学.
|
| [15] |
苏醒, 邹潇潇, 朱军, 等, 2017. 不同光强处理对长茎葡萄蕨藻叶绿素荧光特性的影响[J]. 中国水产科学, 24(4): 783-790.
|
| [16] |
吴启藩, 2017. 不同LED光源对长茎葡萄蕨藻生长、生理生化特性及品质的影响[D]. 湛江: 广东海洋大学.
|
| [17] |
杨宇峰, 2016. 近海环境生态修复与大型海藻资源利用[M]. 北京: 科学出版社:1-364.
|
| [18] |
姚瑶, 2016. 环境因子对针叶蕨藻(Caulerpa sertularioides)生长及氨氮吸收动力学的影响[D]. 海口: 海南大学.
|
| [19] |
余江, 杨宇峰, 聂湘平, 2007. 大型海藻龙须菜对重金属镉胁迫的响应[J]. 四川大学学报(工程科学版), 39(3): 83-90.
|
| [20] |
张永雨, 张继红, 梁彦韬, 等, 2017. 中国近海养殖环境碳汇形成过程与机制[J]. 中国科学: 地球科学, 47(12): 1414-1424.
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}