
海洋酸化和低氧及其节律性变化对海蜇碟状幼体的影响
|
吴钟启悦(1996—), 男, 江西省萍乡市人, 硕士, 从事海洋生物学研究。email: |
Copy editor: 林强
收稿日期: 2022-01-13
修回日期: 2022-03-17
网络出版日期: 2022-03-25
基金资助
国家自然科学基金(Y911051021)
Effects of diurnal changes of ocean acidification and hypoxia on the ephyrae of Rhopilema esculentum
Copy editor: LIN Qiang
Received date: 2022-01-13
Revised date: 2022-03-17
Online published: 2022-03-25
Supported by
National Natural Science Foundation of China(Y911051021)
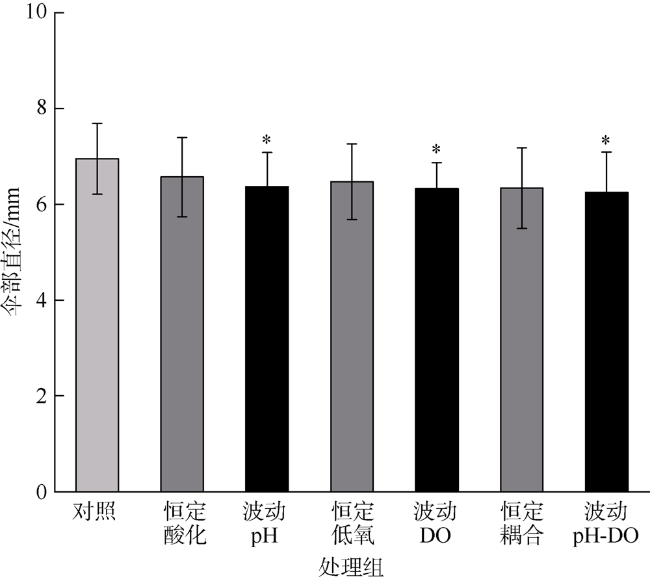
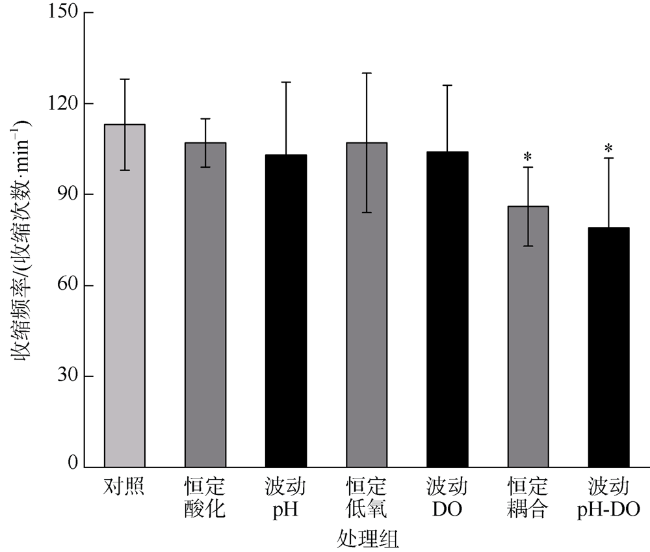
人类生产活动引起的全球气候变化加剧作为重要的环境问题, 海洋中pCO2和溶解氧的失衡及富营养化加剧了许多沿海生态系统的酸化和低氧现象。海蜇(Rhopilema esculentum)为我国重要的渔业资源之一, 具有食用价值及潜在药用价值。本研究评估了海蜇碟状幼体在海洋酸化和低氧及其昼夜节律变化胁迫下的生理响应: 碟状幼体暴露于两个pH水平(酸化pH 7.6, 正常pH 8.1)和两个溶解氧水平(低氧2mg·L-1, 常氧7mg·L-1)维持7d, 胁迫因素维持恒定或昼夜节律性变化, 测定其伞部直径、收缩频率及酸碱平衡、免疫和抗氧化相关酶活力。研究表明, 海洋酸化和低氧在不同程度上影响海蜇碟状幼体的生理代谢和生长发育, 且酸化和低氧之间表现出部分拮抗作用, 昼夜波动的暴露模式对海蜇碟状幼体的损伤程度大于恒定的暴露模式。
吴钟启悦 , 王雷 , 陈立飞 , 李秀保 , 董志军 . 海洋酸化和低氧及其节律性变化对海蜇碟状幼体的影响[J]. 热带海洋学报, 2022 , 41(6) : 114 -124 . DOI: 10.11978/2022007
The anthropogenic aggravation of global climate change is an important environmental problem, and the imbalance between pCO2 and dissolved oxygen in the ocean and eutrophication have aggravated the acidification and hypoxia of many coastal ecosystems. Rhopilema esculentum is one of the most important fishery resources in China, which has edible value and potential medicinal value. In this study, the physiological responses of the ephyrae of R. esculentum under the stress of ocean acidification and hypoxia and their circadian rhythm changes were evaluated: the ephyrae was exposed to two levels of pH (acidified pH 7.6; normal pH 8.1) and two dissolved oxygen (DO) levels (hypoxia 2 mg·L-1; normoxic 7 mg·L-1) for 7 days. The stress factors remained constant or fluctuating, and the bell diameter, pulsation rates and activities of acid-base balance, immunity and antioxidation related enzymes of ephyrae were measured. The results showed that the ocean acidification and hypoxia affected the physiological metabolism, growth and development of R. esculentum ephyrae in different degrees, and there was a partial antagonism between acidification and hypoxia. The effects of diurnal fluctuating exposure mode to R. esculentum ephyrae was more harmful than that of constant exposure mode.
表1 本实验中相关海水化学参数Tab. 1 Chemical parameters of seawater in this experiment (mean±SD) |
| 参数 | 恒定 | 波动pH | 波动DO | 波动pH-DO | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 对照 | 酸化 | 低氧 | 酸化低氧耦合 | 白天 | 夜间 | 平均值 | 白天 | 夜间 | 平均值 | 白天 | 夜间 | 平均值 | ||
| pH | 8.11 ±0.02 | 7.58 ±0.01 | 8.13 ±0.01 | 7.61 ±0.02 | 8.02 ±0.01 | 7.20 ±0.02 | 7.61 ±0.41 | 8.08 ±0.02 | 8.10 ±0.01 | 8.09 ±0.02 | 8.01 ±0.01 | 7.20 ±0.01 | 7.60 ±0.41 | |
| DO/(mg·L-1) | 7.18 ±0.09 | 7.22 ±0.10 | 2.10 ±0.11 | 1.98 ±0.10 | 7.19 ±0.05 | 7.20 ±0.06 | 7.20 ±0.06 | 3.02 ±0.23 | 1.18 ±0.09 | 2.10 ±0.94 | 3.04 ±0.16 | 0.89 ±0.05 | 1.97 ±1.08 | |
| 盐度/‰ | 32.05 ±0.04 | 32.02 ±0.03 | 32.05 ±0.04 | 31.92 ±0.13 | 31.88 ±0.16 | 31.89 ±0.08 | 31.88 ±0.13 | 31.97 ±0.10 | 31.92 ±0.09 | 31.95 ±0.10 | 31.97 ±0.09 | 31.88 ±0.03 | 31.92 ±0.08 | |
| 温度/℃ | 17.97 ±0.15 | 18.04 ±0.03 | 18.13 ±0.16 | 18.04 ±0.12 | 18.03 ±0.12 | 17.97 ±0.16 | 18.00 ±0.14 | 18.17 ±0.08 | 18.09 ±0.08 | 18.13 ±0.09 | 18.11 ±0.18 | 17.91 ±0.07 | 18.01 ±0.17 | |
| TA/(µmol·kg-1) | 2180 ±43 | 2116 ±30 | 2125 ±27 | 2118 ±25 | 2238 ±43 | 2196 ±40 | 2217 ±42 | 2211 ±47 | 2233 ±25 | 2222 ±38 | 2232 ±27 | 2217 ±18 | 2225 ±21 | |
| pCO2/μatm | 463 ±23 | 1679 ±57 | 423 ±14 | 1568 ±72 | 593 ±11 | 4393 ±186 | 2493 ±1904 | 502 ±29 | 480 ±12 | 491 ±25 | 613 ±19 | 4443 ±137 | 2528 ±1917 | |
| DIC/(µmol·kg-1) | 1994 ±41 | 2108 ±28 | 1931 ±26 | 2102 ±25 | 2086 ±34 | 2320 ±35 | 2203 ±122 | 2035 ±47 | 2047 ±17 | 2041 ±36 | 2085 ±24 | 2344 ±13 | 2215 ±131 | |
| 方解石饱和度 | 3.34 ±0.13 | 1.09 ±0.02 | 3.42 ±0.06 | 1.16 ±0.04 | 2.90 ±0.09 | 0.47 ±0.02 | 1.69 ±1.22 | 3.24 ±0.12 | 3.40 ±0.10 | 3.32 ±0.14 | 2.82 ±0.10 | 0.48 ±0.01 | 1.65 ±1.17 | |
| 文石饱和度 | 3.34 ±0.13 | 0.70 ±0.01 | 2.20 ±0.04 | 0.75 ±0.03 | 1.86 ±0.06 | 0.31 ±0.01 | 1.09 ±0.78 | 2.08 ±0.07 | 2.19 ±0.07 | 2.14 ±0.09 | 1.82 ±0.07 | 0.31 ±0.00 | 1.06 ±0.76 | |
表2 双因素方差分析结果: 恒定酸化和低氧胁迫对海蜇碟状幼体的影响Tab. 2 Two-way ANOVA summary: Effects of constant acidification and hypoxia on the R. esculentum ephyrae |
| 生理指标 | 因素 | df | MS | F | P |
|---|---|---|---|---|---|
| 伞部直径 | pH | 1 | 0.980 | 1.470 | 0.231 |
| DO | 1 | 1.912 | 2.865 | 0.096 | |
| pH×DO | 1 | 0.250 | 0.374 | 0.543 | |
| 收缩频率 | pH | 1 | 2522.017 | 10.344 | 0.002 |
| DO | 1 | 2706.817 | 11.102 | 0.002 | |
| pH×DO | 1 | 799.350 | 3.278 | 0.076 | |
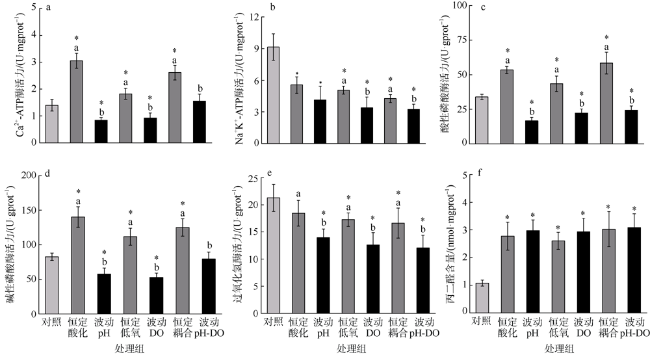
| Ca2+-ATP酶 | pH | 1 | 8.991 | 148.768 | <0.001 |
| DO | 1 | 0.000 | 0.002 | 0.961 | |
| pH×DO | 1 | 1.126 | 18.631 | <0.001 | |
| Na+K+-ATP酶 | pH | 1 | 29.029 | 47.032 | <0.001 |
| DO | 1 | 43.547 | 70.554 | <0.001 | |
| pH×DO | 1 | 11.776 | 19.079 | <0.001 | |
| 酸性磷酸酶 | pH | 1 | 1779.697 | 69.081 | <0.001 |
| 生理指标 | 因素 | df | MS | F | P |
| DO | 1 | 321.720 | 12.488 | 0.002 | |
| pH×DO | 1 | 31.160 | 1.209 | 0.284 | |
| 碱性磷酸酶 | pH | 1 | 7529.052 | 53.969 | <0.001 |
| DO | 1 | 285.784 | 2.049 | 0.168 | |
| pH×DO | 1 | 2938.528 | 21.064 | <0.001 | |
| 过氧化氢酶 | pH | 1 | 17.995 | 3.434 | 0.079 |
| DO | 1 | 51.215 | 9.773 | 0.005 | |
| pH×DO | 1 | 7.190 | 1.372 | 0.255 | |
| 丙二醛 | pH | 1 | 6.736 | 35.227 | <0.001 |
| DO | 1 | 4.767 | 24.934 | <0.001 | |
| pH×DO | 1 | 2.423 | 12.671 | 0.002 |
续表 |
表3 双因素方差分析结果: 波动酸化和低氧胁迫对海蜇碟状幼体的影响Tab. 3 Two-way ANOVA summary: Effects of fluctuating acidification and hypoxia on the R. esculentum ephyrae |
| 生理指标 | 因素 | df | MS | F | P |
|---|---|---|---|---|---|
| 伞部直径 | pH | 1 | 1.653 | 3.065 | 0.085 |
| DO | 1 | 2.046 | 3.793 | 0.056 | |
| pH×DO | 1 | 0.968 | 1.794 | 0.186 | |
| 收缩频率 | pH | 1 | 4437.600 | 9.869 | 0.003 |
| DO | 1 | 4001.667 | 8.900 | 0.004 | |
| pH×DO | 1 | 912.600 | 2.030 | 0.160 | |
| Ca2+-ATP酶 | pH | 1 | 0.007 | 0.177 | 0.678 |
| DO | 1 | 0.093 | 2.329 | 0.143 | |
| pH×DO | 1 | 2.097 | 52.710 | <0.001 | |
| Na+K+-ATP酶 | pH | 1 | 40.397 | 35.235 | <0.001 |
| DO | 1 | 66.475 | 57.982 | <0.001 | |
| pH×DO | 1 | 35.541 | 31.000 | <0.001 | |
| 酸性磷酸酶 | pH | 1 | 343.219 | 47.102 | <0.001 |
| DO | 1 | 24.000 | 3.294 | 0.085 | |
| pH×DO | 1 | 558.655 | 76.668 | <0.001 | |
| 碱性磷酸酶 | pH | 1 | 4.621 | 0.075 | 0.787 |
| DO | 1 | 95.914 | 1.563 | 0.226 | |
| pH×DO | 1 | 4052.673 | 66.045 | <0.001 | |
| 过氧化氢酶 | pH | 1 | 94.056 | 19.397 | <0.001 |
| DO | 1 | 167.074 | 34.456 | <0.001 | |
| pH×DO | 1 | 68.204 | 14.066 | 0.001 | |
| 丙二醛 | pH | 1 | 6.288 | 39.084 | <0.001 |
| DO | 1 | 5.818 | 36.164 | <0.001 | |
| pH×DO | 1 | 4.597 | 28.572 | <0.001 |
图3 海蜇碟状幼体暴露于恒定/波动的酸化和/或低氧处理后的酶活力*表示和对照组有显著差异(P<0.05); 误差棒上方不同英文小写字母表示恒定和波动的实验组之间存在显著差异(P<0.05) Fig. 3 Enzyme activities of R. esculentum ephyrae after constant/fluctuating acidification and/or hypoxia exposure. The * above error bar indicates the significant difference (P<0.05) as compared to control group; the different lowercase letters indicate significant differences (P<0.05) between constant and fluctuating treatment groups |
| [1] |
丁耕芜, 陈介康, 1981. 海蜇的生活史[J]. 水产学报, 5(2): 93-102.
|
| [2] |
韩春艳, 何晓琳, 李安文, 等, 2016. 酸碱胁迫对奥尼罗非鱼幼鱼鳃组织结构及Na+-K+-ATP酶活力的影响[J]. 安徽农业科学, 44(11): 273-276.
|
| [3] |
霍达, 2020. 刺参应对高温低氧胁迫的生理响应与分子调控特征[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 1-183.
|
| [4] |
姜娓娓, 方建光, 李加琦, 等, 2017. 温度胁迫对皱纹盘鲍生理和生化活动的影响[J]. 中国水产科学, 24(2): 220-230.
|
| [5] |
李飞, 石加慧, 臧瑜, 等, 2018. 海洋酸化对日本虎斑猛水蚤发育、繁殖以及ATP酶活性的影响[J]. 海洋湖沼通报, (4): 44-50.
|
| [6] |
李云峰, 李玉龙, 周遵春, 等, 2020. 我国北方地区海蜇池塘养殖技术研究进展[J]. 水产科学, 39(2): 286-291.
|
| [7] |
刘顶海, 2011. 海蜇(Rhopilema esculenta)幼体发育、苗种培育及生长规律的研究[D]. 上海: 上海海洋大学: 1-65.
|
| [8] |
刘辉, 赵建民,王清, 2018-07-03. 一种海洋酸化和低氧模拟装置及其控制方法: 中国, CN108241394A[P].
|
| [9] |
宋金明, 王启栋, 2021. 近40年来对南海化学海洋学研究的新认知[J]. 热带海洋学报, 40(3): 15-24.
|
| [10] |
宋晶, 吴垠, 李晓东, 等, 2009. 海蜇生长、存活影响因子的研究现状[J]. 河北渔业, (6): 45-49.
|
| [11] |
孙婷婷, 刘中援, 梁丽琨, 等, 2017. 海蜇幼体环介导等温扩增快速检测方法的建立和应用[J]. 应用海洋学学报, 36(4): 538-544.
|
| [12] |
王国善, 于志刚, 米铁柱, 等, 2014. 环境因子对海蜇生长发育影响的研究进展[J]. 海洋科学, 38(1): 85-90.
|
| [13] |
王雷, 2020. 海月水母对海洋酸化和铜离子胁迫的生理响应研究[D]. 烟台: 中国科学院大学(中国科学院烟台海岸带研究所): 1-79.
|
| [14] |
王晓芹, 2018. 海洋酸化胁迫对紫贻贝和长牡蛎生理活动的影响及其缓解途径[D]. 上海: 上海海洋大学: 1-68.
|
| [15] |
韦钦胜, 王保栋, 于志刚, 等, 2017. 夏季长江口外缺氧频发的机制及酸化问题初探[J]. 中国科学: 地球科学, 47(1): 114-134.
|
| [16] |
文春根, 张丽红, 胡宝庆, 等, 2009. pH对背角无齿蚌(Anodonta woodiana)5种免疫因子的影响[J]. 南昌大学学报(理科版), 33(2): 172-176.
|
| [17] |
肖武汉, 2014. 低氧信号传导途径与鱼类低氧适应[J]. 中国科学: 生命科学, 44(12): 1227-1235.
|
| [18] |
徐彦, 2012. 菲律宾蛤仔(Ruditapes philippinarum)对重金属离子—Cd2+、Pb2+免疫应激响应的研究[D]. 青岛: 中国海洋大学: 1-83.
|
| [19] |
曾姣, 2021. 大黄鱼幼鱼对酸化-低氧胁迫的生理响应[D]. 上海: 上海海洋大学: 1-103.
|
| [20] |
张培玉, 唐学玺, 董双林, 2007. 蒽对太平洋牡蛎不同组织抗氧化酶活性差异性影响与膜脂质过氧化研究[J]. 海洋环境科学, 26(5): 434-437.
|
| [21] |
朱晓闻, 2012. 温度、盐度、pH对马氏珠母贝稚贝生理学特征的联合效应研究[D]. 湛江: 广东海洋大学: 1-98.
|
| [22] |
庄淑芳, 张民, 陈芳奕, 2014. 海洋生物响应海洋酸化研究进展[J]. 海峡科学, (8): 3-6. (in Chinese)
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}