
基于ZooScan图像分析“海马”冷泉区浮游动物垂直分布特征
|
李若飞(1998—), 男, 研究方向为海洋浮游生态学。email: |
Copy editor: 殷波
收稿日期: 2022-03-24
修回日期: 2022-05-01
网络出版日期: 2022-05-05
基金资助
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0401)
国家自然科学基金项目(32171548)
国家自然科学基金项目(31971432)
国家自然科学基金项目(41976112)
国家科技基础资源调查专项(2017FY201404)
Vertical distribution of zooplankton in the “Haima” cold seep region based on ZooScan image analysis
Copy editor: YIN Bo
Received date: 2022-03-24
Revised date: 2022-05-01
Online published: 2022-05-05
Supported by
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0401)
National Natural Science Foundation of China(32171548)
National Natural Science Foundation of China(31971432)
National Natural Science Foundation of China(41976112)
Science and Technology Basic Resources Investigation Program of China(2017FY201404)
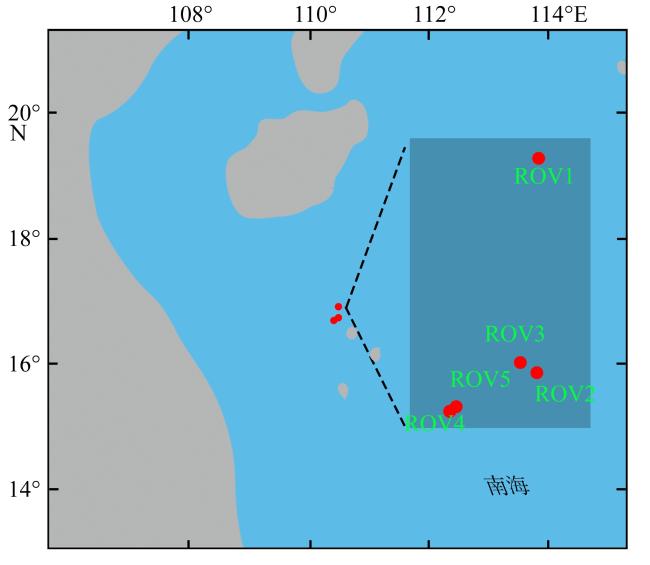
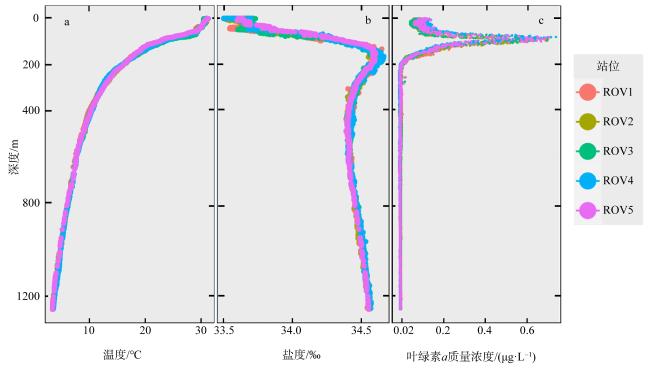
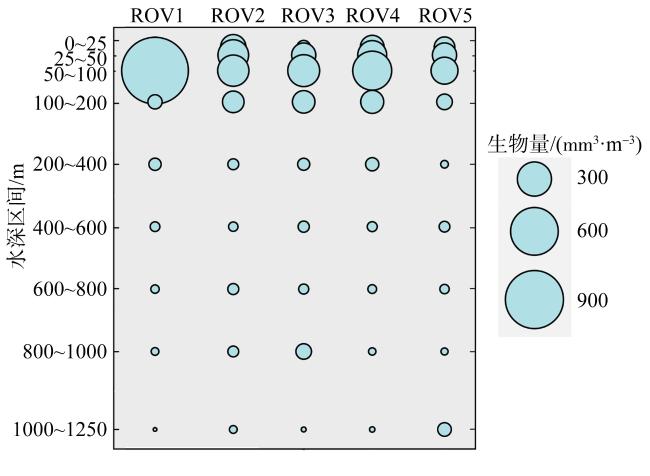
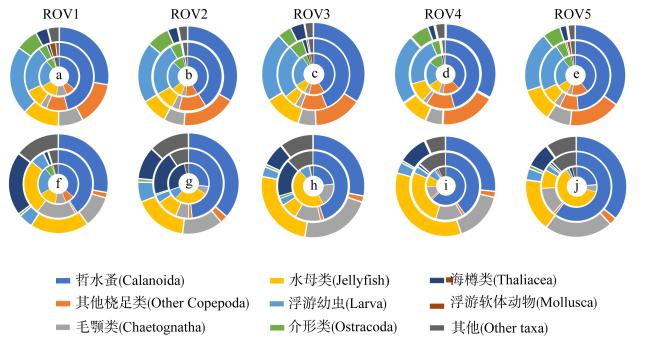
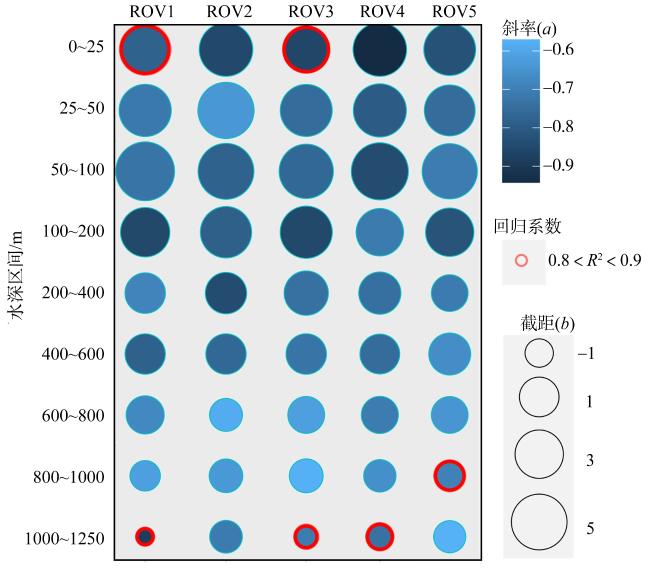
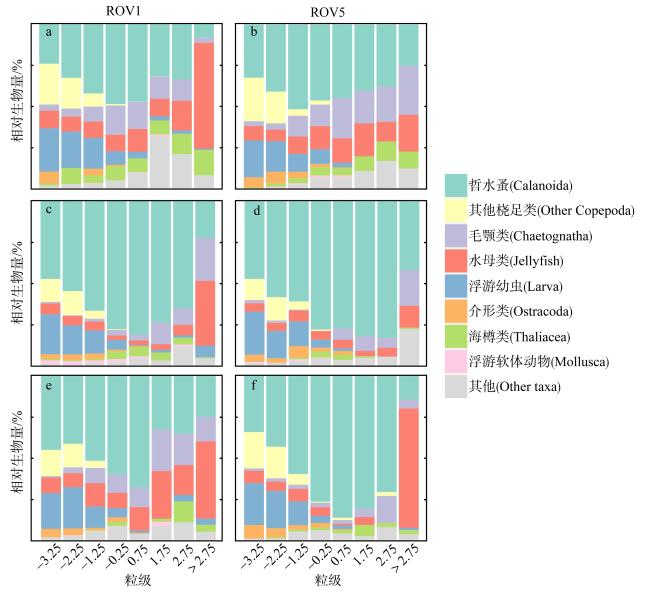
为了解“海马”冷泉区浮游动物的垂直分布以及冷泉活动可能对上方浮游动物群落产生的影响, 2020年9月在“海马”冷泉区的5个调查站位使用浮游生物分层拖网对1250m以浅的浮游动物垂直分布进行了调查。使用浮游动物图像扫描系统(ZooScan)对不同水层的浮游动物样品进行了分析, 获取了浮游动物的丰度、生物量和粒径谱的垂直分布特征。结果显示: 浮游动物丰度和生物量主要集中在0 ~ 100m水层, 低于100m后, 浮游动物的丰度和生物量随着水深增加均快速下降, 在1000 ~ 1250m水层平均分别仅为8.33个·m-3和12.10mm3·m-3。 总体看来, 桡足类是各水层浮游动物的优势类群, 胶质类浮游动物在深层中的占比上升。“海马”冷泉区不同站位和水层标准化生物量谱斜率的变动范围为-0.94 ~ -0.57, 截距的变化范围为-2.10 ~ 5.94。从表层到底层, 浮游动物标准化生物量谱基本呈现斜率逐渐增大而截距逐渐减小的趋势。这反映了浮游生态系统的生产力水平从表到底逐渐下降, 但浮游食物网的能量传递效率逐渐增加。甲烷气体渗漏强度最大的ROV1站位在1000 ~ 1250m水层表现出异常的粒径谱特征, 生物量谱的斜率a显著低于同水层其他站位, 同时在600 ~ 800m水层出现浮游动物丰度的相对高值, 推测该站位的浮游动物群落结构可能受到冷泉区甲烷渗漏的影响。
李若飞 , 柯志新 , 李开枝 , 刘甲星 , 谭烨辉 . 基于ZooScan图像分析“海马”冷泉区浮游动物垂直分布特征[J]. 热带海洋学报, 2023 , 42(2) : 87 -96 . DOI: 10.11978/2022057
To understand the vertical distributions of zooplankton in regions of the “Haima” cold seep and the possible effects of seep activities, zooplankton samples were collected from 0 to 1250 m at five stations in September 2020. These zooplankton samples were analyzed using ZooScan image analysis system. The abundance, biovolume and size spectra of zooplankton in different water layers were obtained in this study. The results showed that zooplankton abundance and biovolume were mainly occurred in the 0 ~ 100 m water layer. Below 100 m, zooplankton abundance and biovolume decreased rapidly with increased depth. The average abundance and biovolume of zooplankton were only 8.33 ind·m-3 and 12.10 mm3·m-3 at the layer of 1000 ~ 1250 m. In general, copepods were the dominant zooplankton in each water layer, and the proportion of gelatinous zooplankton increased in deep layer. The slope of normalized biovolume size spectra (NBSS) in the regions of the “Haima” cold seep ranged from -0.94 to -0.57, and the intercept ranged from -2.10 to 5.94. From surface to bottom, the slope of NBSS increased while the intercept decreased gradually. This indicated that the productivity of planktonic ecosystem declined gradually from surface to bottom, but the energy transfer efficiency of pelagic food web increased gradually. The layer of 1000 ~ 1250 m at ROV1 station showed abnormal size spectrum characteristics, and the slope “a” is significantly lower than that of other stations in the same water layer. It suggested that the zooplankton community might be affected by cold seep activities.
图3 “海马”冷泉0 ~ 1250m 浮游动物总丰度的垂直分布图Fig. 3 Vertical distributions of total zooplankton abundance (ind·m-3) from surface to 1250 m at “Haima” cold seeps |
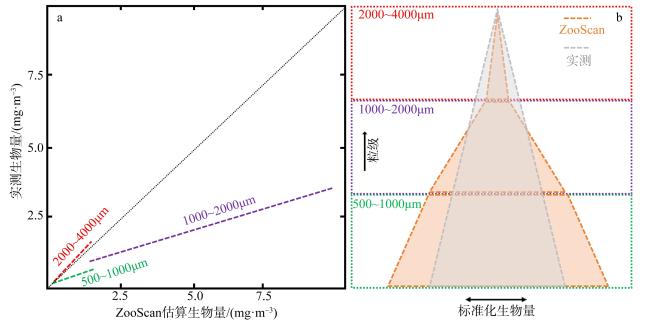
图8 基于ZooScan估算生物量并描述浮游动物粒级分布与实际测量的差异示意图a. 不同粒级ZooScan测量生物量与实测生物量的线性关系[修改自Koplin (2020)]; b. 不同的粒级ZooScan计算和实际测量的生物量的差异。标准化生物量代表生物量除以粒级间隔 Fig. 8 Schematic diagram showing differences between Zooscan image analysis and manual measurements in biomass calculation and in describing zooplankton size distribution. (a) the linear relationship between ZooScan measured biomass and manually measured biomass, revised from Koplin (2020); (b) the size pyramid based on this linear relationship. Standardized biomass (or biovolume) represents biomass/size interval |
| [1] |
代鲁平, 李超伦, 王世伟, 等, 2016. 基于ZooScan图像技术的南黄海夏季浮游动物群落结构分析[J]. 海洋与湖沼, 47(4): 764-773.
|
| [2] |
柯志新, 黄良民, 谭烨辉, 等, 2013. 2008年夏末南海北部叶绿素a的空间分布特征及其影响因素[J]. 热带海洋学报, 32(4): 51-57.
|
| [3] |
李开枝, 任玉正, 柯志新, 等, 2021. 南海东北部陆坡区中上层浮游动物的垂直分布[J]. 热带海洋学报, 40(2): 61-73.
|
| [4] |
连喜平, 谭烨辉, 刘永宏, 等, 2013. 两种浮游生物网对南海北部浮游动物捕获效率的比较[J]. 热带海洋学报, 32(3): 33-39.
|
| [5] |
孙松, 毕永坤, 孙晓霞, 2013. 基于图像技术的胶州湾浮游动物优势种体型参数与生物量转换关系研究[J]. 海洋与湖沼, 44(1): 15-22.
|
| [6] |
赵静, 龚跃华, 何雪宝, 等, 2020. 南海北部陆坡西部海域“海马”冷泉甲烷渗漏及其海底表征[J]. 地球化学, 49(1): 108-118.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}