
海南岛岸礁澄黄滨珊瑚(Porites lutea)集合种群的遗传结构和连通性
|
付成冲(1991—), 男, 山东省菏泽市人, 硕士研究生, 从事珊瑚生态遗传学研究。email: |
Copy editor: 殷波
收稿日期: 2022-05-04
修回日期: 2022-05-13
网络出版日期: 2022-05-16
基金资助
国家自然科学基金项目(41376174)
海南省重点研发计划项目(ZDYF2018108)
国家重点研发计划项目(2018YFC1406504)
The genetic structure and connectivity of Porites lutea metapopulation of the fringing reefs around the Hainan Island
Copy editor: YIN Bo
Received date: 2022-05-04
Revised date: 2022-05-13
Online published: 2022-05-16
Supported by
National Natural Science Foundation of China(41376174)
Key Research and Development Program of Hainan Province(ZDYF2018108)
National Key Research and Development Program of China(2018YFC1406504)
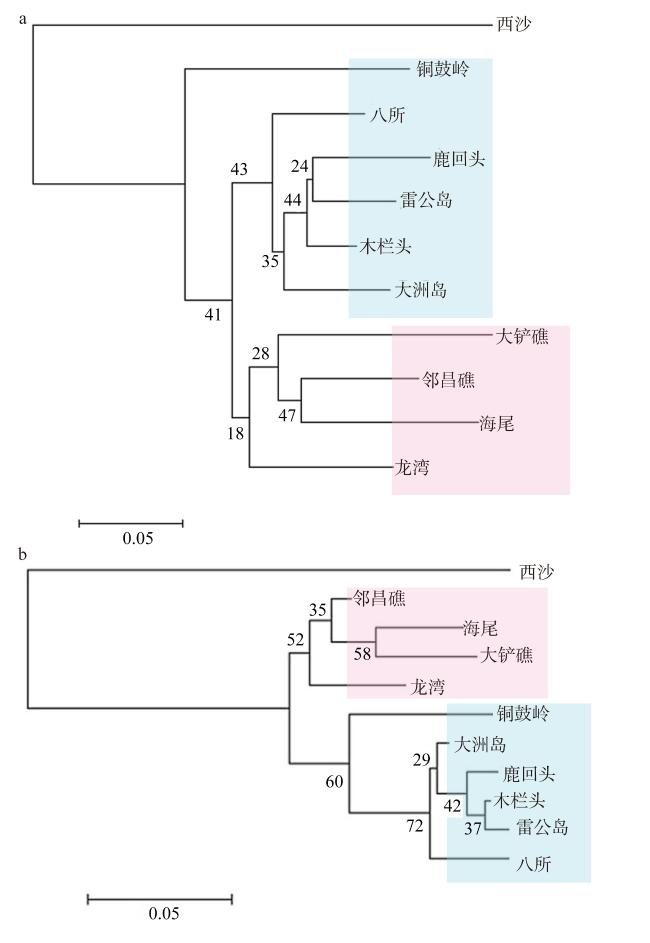
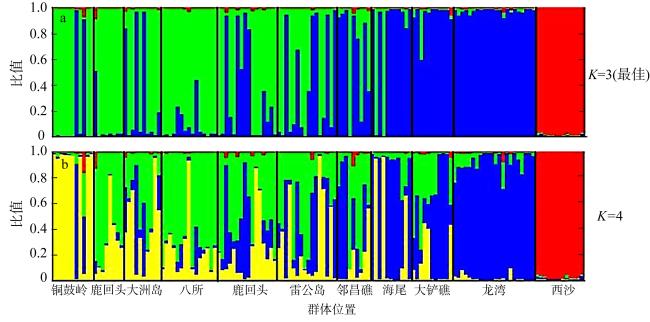
海南岛岸礁的造礁珊瑚代表性种类澄黄滨珊瑚(Porites lutea)是环境适应性较强的块状产卵型珊瑚。探究其遗传结构和连通性将有助于揭示其海南岛岸礁集合种群的遗传多样性格局和幼虫迁移路径, 进而阐明海南珊瑚礁的恢复潜力。文章通过11个P. lutea微卫星标记来分析10个海南岛岸礁地理群体(八所、海尾、大铲礁、邻昌礁、雷公岛、木栏头、铜鼓岭、龙湾、大洲岛和鹿回头)和1个西沙群岛(西沙七连屿)群体的遗传结构。结果显示, 整体上各群体的遗传多样性中等偏低, 平均等位基因丰度 (allelic richness, Rs)为(2.8±1.3)(八所群体) ~ (3.7±1.7)(邻昌礁群体), 平均观测杂合度和期望杂合度分别为0.31 (铜鼓岭群体) ~ 0.54 (大铲礁群体)和0.50 (雷公岛群体) ~ 0.64 (海尾群体)。除海南岛东部龙湾群体、大洲岛群体和西部的八所群体、大铲礁群体之外, 其他7/11的地理群体均呈杂合子缺失。海南岛岸礁澄黄滨珊瑚P. lutea地理群体间的遗传分化显示, 集合种群分为北东南遗传连通带和西岸两支, 支间遗传分化显著, 前者包括北岸的雷公岛群体和木栏头群体、东岸的大洲岛群体, 以及南岸的鹿回头和八所群体。由于珊瑚幼虫随海流迁移而形成的有效的基因流, 消弭了它们之间的遗传分化, 而西岸群体因北部湾沿岸海流交换不畅, 与外部的基因交流受阻。西岸的海尾群体与西南部的八所群体间尽管相距不足50km, 但遗传分化明显, 这可能是由于昌化江径流形成的盐度波动和悬浮沉积物的隔离作用。同样, 铜鼓岭群体因处于铜鼓岭岬角内波影区, 其南部八门湾径流或限制了它与东岸群体的基因交流, 因而呈现近交、低杂合度和非随机交配特性。此外, 由于对离岸礁坡环境的趋同适应, 东岸龙湾群体与西岸的离岸岛礁邻昌礁群体、大铲礁群体和海尾群体之间的遗传相似性更高。以P. lutea集合种群为代表的海南岛珊瑚岸礁有着相对充沛的基因交流, 且因受海流、径流及复杂岸礁环境影响而形成了多层次的遗传分化和基因流格局, 这构成其应对外界胁迫的主要自然恢复力。西沙群体与海南岛岸礁群体之间的遗传分化较大, 呈现显著的地理隔离。西沙七连屿可能由于自身珊瑚礁健康状况的衰退而丧失了对海南岛珊瑚幼虫的补充能力。
付成冲 , 李福宇 , 陈丹丹 , 侯敬 , 王珺 , 李元超 , 王道儒 , 王嫣 . 海南岛岸礁澄黄滨珊瑚(Porites lutea)集合种群的遗传结构和连通性[J]. 热带海洋学报, 2023 , 42(2) : 64 -77 . DOI: 10.11978/2022098
Porites lutea, the representative species of reef building corals around the Hainan Island, is a spawning, massive coral with strong environmental adaptability. Exploring the genetic structure and connectivity of this species helps to reveal the genetic diversity pattern and larval migration path of coral metapopulation around the Hainan Island, thus clarifying the recovery potential of coral reefs. In this study, 11 P. lutea microsatellite markers were screened to analyze the genetic structure of 10 populations of the Hainan fringing reefs and 1 population (XsR) in the Xisha Islands. The results showed that, overall the genetic diversity of all populations was medium to low, with the average allelic richness Rs ranging from 2.8 ± 1.3 (Basuo population, Bs) to 3.7 ± 1.7 (Linchang Reef population, LcR), and the average observed and expected heterozygosity ranged from 0.31 (Tongguling population, Tgl) to 0.54 (Dachan Reef population, DcR) and 0.50 (Leigong Island population, LgI) to 0.64 (Haiwei population, Hw), respectively. Except for the Longwan Reef population (LwR) and the Dazhou Island population (DzI), which located in the east of the Hainan Island, and the Basuo population (Bs) and the Dachan Reef population DcR (in the west of Hainan Island), all other populations (7/11) showed evidence of heterozygote deficiency. According to genetic differentiation, the Hainan Island populations were divided into two groups: the north-south-east genetically connected zone and the west coast, and the differentiation between the two branches (AMOVA, 0.092) was significant. The former group included Bs, Luhuitou population (Lht), DzI, LgI and Mulantou population (Mlt), due to the significant gene flow created by exchange of ocean currents, there was no obvious genetic differentiation among these five coastal populations, whereas the gene flow of the offshore populations (LcR, DcR and Hw) of the west coast was blocked due to the discontinuity of coastal reefs and slow coastal currents. LwR in the east coast also converges to the west branch, which may be due to the isolation by environment and the convergent adaptation to offshore environment. Although the distance between Hw and Bs is less than 50 km, but they are obviously differentiated, possibly due to the isolation by salinity fluctuation and suspended sediments caused by the runoff of the Changhua River. Tgl showed strong inbreeding, low heterozygosity and non-random mating characteristics because it was located in the wave shadow area of the Tongguling headland, and the runoff of the Bamen River restricted its gene exchange with other coastal reef populations. Represented by P. lutea metapopulations, the fringing reefs of the Hainan Island has the natural resilience responding to environmental stress due to the strong gene flow and the genetic differentiation caused by ocean current, runoff and complex fringing reef structure, as well as the environmental differences between nearshore and offshore. The genetic differentiation between the Xisha Islands population and the coastal reef populations of the Hainan Island was large, showing significant geographical isolation. The Qilianyu Island may have lost the ability to replenish the Hainan Island with coral larvae because the coral reefs have declined there.
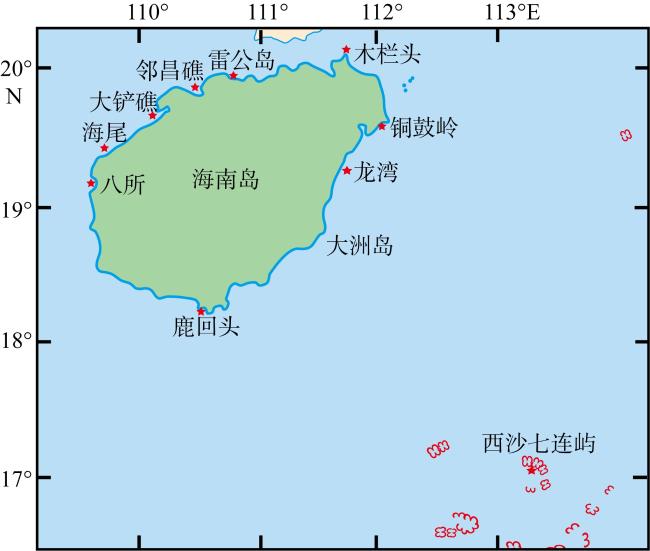
图1 采样点地图该图基于国家测绘地理信息局标准地图服务网站下载的审图号为琼S(2021)121的标准地图制作, 底图无修改 Fig. 1 Map of sampling sites |
表1 澄黄滨珊瑚样品信息表Tab. 1 Porites lutea samples details |
| 地区 | 站位 | 纬度/N | 经度/E | N | NMLG |
|---|---|---|---|---|---|
| 海 南 岛 岸 礁 | 八所 | 19°08′25.80″ | 108°39′33.00″ | 15 | 15 |
| 雷公岛 | 19°58′30.43″ | 109°53′29.79″ | 17 | 16 | |
| 木栏头 | 20°07′25.02″ | 110°39′21.90″ | 16 | 16 | |
| 铜鼓岭 | 19°37′53.39″ | 110°58′42.76″ | 17 | 11 | |
| 大洲岛 | 18°40′29.54″ | 110°28′56.86″ | 10 | 10 | |
| 鹿回头 | 18°13′33.42″ | 109°29′06.44″ | 8 | 8 | |
| 海尾 | 19°26′47.55″ | 108°51′12.20″ | 10 | 10 | |
| 大铲礁 | 19°41′11.63″ | 109°06′2.13″ | 11 | 11 | |
| 邻昌礁 | 19°55′01.09″ | 109°28′51.86″ | 10 | 9 | |
| 龙湾 | 19°16′24.20″ | 110°39′29.50″ | 22 | 22 | |
| 西沙岛礁 | 七连屿 | 16°58′10.92″ | 112°18′21.41″ | 13 | 13 |
| 总计 | 149 | 141 |
注: N为正常扩增的样品数; NMLG为去除遗传同质性克隆个体后的样品数 |
表2 11个微卫星位点在11个澄黄滨珊瑚群体的扩增条件和遗传信息(李福宇 等, 2021)Tab. 2 Information and characteristics of 11 microsatellite loci in total 141 wild P. lutea colonies from 11 populations |
| 位点 | GenBank号 | Ta/℃ | a | Size/bp | Ho | RS | HWE* | PIC |
|---|---|---|---|---|---|---|---|---|
| plo7 | HQ435873 | 50 | 12 | 171~240 | 0.44 ± 0.21 | 4.2 ± 1.2 | 1/10 # | 0.762 |
| plo37 | HQ435922 | 55 | 17 | 372~431 | 0.56 ± 0.12 | 5.4 ± 0.9 | 2/11 | 0.824 |
| plo66 | HQ435976 | 55 | 11 | 172~208 | 0.36 ± 0.22 | 2.9 ± 1.0 | 0/10 # | 0.443 |
| plo78 | GU137158 | 50 | 9 | 102~170 | 0.52 ± 0.27 | 2.9 ± 1.0 | 2/9 # | 0.640 |
| pasE005 | KP407156 | 53 | 2 | 202~204 | 0.16 ± 0.15 | 1.6 ± 0.4 | 0/5 # | 0.150 |
| pasE030 | KP407159 | 53 | 4 | 183~197 | 0.18 ± 0.11 | 1.5 ± 0.5 | 0/5 # | 0.093 |
| pasE056 | KP407161 | 50 | 8 | 251~300 | 0.25 ± 0.15 | 2.7 ± 0.6 | 1/11 | 0.432 |
| pasE060 | KP407163 | 50 | 18 | 261~392 | 0.64 ± 0.15 | 5.9 ± 0.9 | 3/11 | 0.850 |
| pasE062 | KP407165 | 53 | 5 | 288~314 | 0.23 ± 0.14 | 2.4 ± 0.5 | 0/11 | 0.309 |
| pasE073 | KP407169 | 48 | 9 | 326~357 | 0.73 ± 0.15 | 3.3 ± 0.6 | 2/11 | 0.600 |
| pasE099 | KP407171 | 53 | 6 | 202~302 | 0.38 ± 0.35 | 2.0 ±0.8 | 0/6 # | 0.350 |
注: Ta为退火温度; a为等位基因数; Size为片段大小, Ho为观测杂合度; RS为等位基因丰度; HWE*为显著偏离哈迪-温伯格平衡的群体数/检测群体数(α=0.05/11=0.0045); #表示检测群体数少于11 (总群体数), 因有的群体由于等位基因数太少而不能进行哈迪温伯格平衡检测; PIC为多态信息含量, 基于全部141个个体的基因型计算 |
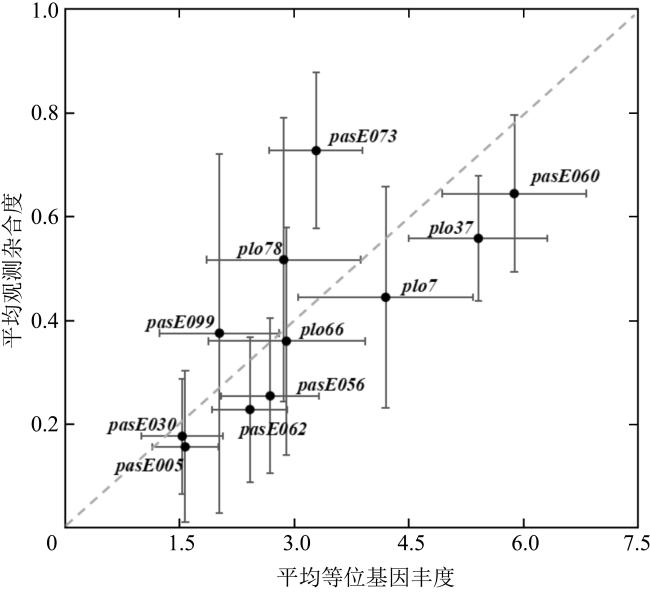
图2 11个微卫星位点在11个澄黄滨珊瑚地理群体中所呈现的遗传多样性每个黑点的水平和垂直线段表示标准差 Fig. 2 Diversity of 11 microsatellite sites in 141 individuals of 11 geographic populations of P. lutea. Each black dot represents the mean allelic richness Rs (horizontal axis) and the mean observed heterozygous Ho (vertical axis) for a microsatellite locus. The length of horizontal and vertical line represents the standard deviations |
表3 11个澄黄滨珊瑚群体11个微卫星位点的遗传多样性参数表Tab. 3 Summary statistics for 11 microsatellite loci among 11 Porites lutea populations |
| 群体 | 指标 | 位点 | 平均值 | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| plo7 | plo37 | plo66 | plo78 | pasE005 | pasE030 | pasE056 | pasE060 | pasE062 | pasE073 | pasE099 | |||||||||||||||||||||||||||||
| FST | 0.223 | 0.093 | 0.122 | 0.285 | 0.087 | 0.033 | 0.064 | 0.012 | 0.084 | 0.095 | 0.303 | ||||||||||||||||||||||||||||
| 铜 鼓 岭 | N | 9 | 10 | 10 | 11 | 10 | 11 | 11 | 11 | 11 | 11 | 10 | |||||||||||||||||||||||||||
| RS | 4.0 | 3.8 | 2.9 | 2.6 | 2.0 | 1.9 | 2.0 | 5.6 | 1.9 | 3.3 | 2.7 | 3.0 | |||||||||||||||||||||||||||
| Ho | 0.27 | 0.55 | 0.18 | 0.09 | 0.09 | 0.18 | 0.00 | 0.91 | 0.00 | 0.91 | 0.27 | 0.31 | |||||||||||||||||||||||||||
| He | 0.85 | 0.74 | 0.67 | 0.62 | 0.47 | 0.26 | 0.61 | 0.82 | 0.26 | 0.61 | 0.59 | 0.59 | |||||||||||||||||||||||||||
| FIS | 0.556 | 0.115 | 0.660 | 0.839 | 0.640 | -0.053 | 1.000 | -0.117 | 1.000 | -0.538 | 0.289 | 0.399 | |||||||||||||||||||||||||||
| Phwe | 0.0075* | 0.0020 | 0.0056* | 0.0011* | 0.1603 | 1.0000 | 0.0007* | 0.0006 | 0.0490 | 0.0037 | 0.4864 | ||||||||||||||||||||||||||||
| 鹿 回 头 | N | 8 | 8 | 7 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 7 | |||||||||||||||||||||||||||
| RS | 5.0 | 5.6 | 1.0 | 2.9 | 2.0 | 2.0 | 2.0 | 5.7 | 3.0 | 3.8 | 2.0 | 3.2 | |||||||||||||||||||||||||||
| Ho | 0.50 | 0.63 | 0.00 | 0.50 | 0.38 | 0.38 | 0.25 | 0.50 | 0.25 | 0.75 | 0.13 | 0.39 | |||||||||||||||||||||||||||
| He | 0.89 | 0.88 | 0.35 | 0.54 | 0.46 | 0.33 | 0.34 | 0.86 | 0.53 | 0.62 | 0.45 | 0.57 | |||||||||||||||||||||||||||
| FIS | 0.411 | 0.239 | — | 0.082 | 0.192 | -0.167 | -0.077 | 0.404 | 0.440 | -0.235 | 0.000 | 0.129 | |||||||||||||||||||||||||||
| Phwe | 0.0214 | 0.2890 | — | 1.0000 | 1.0000 | 1.0000 | 1.0000 | 0.0176* | 0.1413 | 0.0417 | — | ||||||||||||||||||||||||||||
| 大 洲 岛 | N | 9 | 9 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 9 | 9 | |||||||||||||||||||||||||||
| RS | 5.3 | 7.0 | 3.6 | 4.1 | 1.7 | 1.9 | 1.9 | 5.7 | 1.9 | 2.0 | 1.8 | 3.4 | |||||||||||||||||||||||||||
| Ho | 0.60 | 0.70 | 0.50 | 0.50 | 0.10 | 0.20 | 0.20 | 0.70 | 0.20 | 0.50 | 0.10 | 0.39 | |||||||||||||||||||||||||||
| He | 0.81 | 0.88 | 0.56 | 0.68 | 0.19 | 0.28 | 0.28 | 0.86 | 0.28 | 0.54 | 0.37 | 0.52 | |||||||||||||||||||||||||||
| FIS | 0.127 | 0.097 | 0.100 | 0.244 | 0.000 | -0.059 | -0.059 | 0.187 | -0.059 | -0.333 | 0.000 | 0.022 | |||||||||||||||||||||||||||
| Phwe | 0.1919 | 0.1379 | 0.1023 | 0.5001 | — | 1.0000 | 1.0000 | 0.0600 | 1.0000 | 1.0000 | — | ||||||||||||||||||||||||||||
| 八 所 | N | 15 | 14 | 15 | 15 | 15 | 15 | 14 | 15 | 15 | 14 | 14 | |||||||||||||||||||||||||||
| RS | 3.7 | 4.3 | 2.0 | 2.7 | 2.0 | 1.0 | 2.5 | 5.1 | 2.0 | 3.5 | 1.5 | 2.8 | |||||||||||||||||||||||||||
| Ho | 0.80 | 0.40 | 0.60 | 0.80 | 0.40 | — | 0.27 | 0.53 | 0.40 | 0.80 | 0.07 | 0.51 | |||||||||||||||||||||||||||
| He | 0.72 | 0.76 | 0.54 | 0.54 | 0.40 | — | 0.46 | 0.79 | 0.40 | 0.70 | 0.25 | 0.56 | |||||||||||||||||||||||||||
| FIS | -0.139 | 0.371 | -0.260 | -0.500 | 0.012 | — | 0.096 | 0.311 | 0.012 | -0.316 | 0.000 | -0.041 | |||||||||||||||||||||||||||
| Phwe | 0.5795 | 0.0460 | 0.5809 | 0.0684 | 1.0000 | — | 0.1472 | 0.0115 | 1.0000 | 0.0132 | — | ||||||||||||||||||||||||||||
| 木 栏 头 | N | 16 | 15 | 16 | 15 | 15 | 16 | 16 | 16 | 16 | 15 | 15 | |||||||||||||||||||||||||||
| RS | 5.0 | 5.6 | 2.3 | 3.6 | 1.5 | 2.4 | 2.4 | 4.9 | 2.8 | 3.4 | 1.0 | 3.2 | |||||||||||||||||||||||||||
| Ho | 0.50 | 0.75 | 0.19 | 0.50 | 0.06 | 0.13 | 0.25 | 0.63 | 0.25 | 0.75 | 0.00 | 0.36 | |||||||||||||||||||||||||||
| He | 0.77 | 0.84 | 0.24 | 0.68 | 0.24 | 0.29 | 0.56 | 0.76 | 0.47 | 0.64 | 0.18 | 0.52 | |||||||||||||||||||||||||||
| FIS | 0.324 | 0.003 | -0.034 | 0.161 | 0.000 | 0.474 | 0.514 | 0.169 | 0.409 | -0.388 | — | 0.163 | |||||||||||||||||||||||||||
| Phwe | 0.0182* | 0.9930 | 1.0000 | 0.0524 | — | 0.0675 | 0.0234 | 0.0286 | 0.0683 | 0.1924 | — | ||||||||||||||||||||||||||||
| 雷 公 岛 | N | 16 | 16 | 16 | 16 | 15 | 15 | 16 | 16 | 16 | 16 | 15 | |||||||||||||||||||||||||||
| RS | 5.5 | 5.7 | 1.8 | 4.6 | 1.9 | 1.9 | 3.3 | 6.0 | 1.8 | 2.4 | 1.0 | 3.3 | |||||||||||||||||||||||||||
| Ho | 0.63 | 0.56 | 0.06 | 0.63 | 0.06 | 0.06 | 0.56 | 0.56 | 0.19 | 0.50 | 0.00 | 0.35 | |||||||||||||||||||||||||||
| He | 0.83 | 0.84 | 0.23 | 0.74 | 0.34 | 0.29 | 0.54 | 0.87 | 0.23 | 0.40 | 0.18 | 0.50 | |||||||||||||||||||||||||||
| FIS | 0.254 | 0.322 | 0.651 | 0.143 | 0.650 | 0.500 | -0.034 | 0.348 | -0.071 | -0.257 | — | 0.251 | |||||||||||||||||||||||||||
| Phwe | 0.0189 | 0.0903* | 0.0962 | 0.0042 | 0.1023 | 0.0306 | 0.7697 | 0.0025* | 1.0000 | 0.6317 | — | ||||||||||||||||||||||||||||
| 邻 昌 礁 | N | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | |||||||||||||||||||||||||||
| RS | 4.6 | 6.3 | 3.6 | 1.8 | 2.0 | 1.8 | 3.7 | 6.8 | 3.0 | 4.0 | 3.7 | 3.7 | |||||||||||||||||||||||||||
| Ho | 0.56 | 0.56 | 0.44 | 0.11 | 0.11 | 0.11 | 0.44 | 0.67 | 0.22 | 1.00 | 0.78 | 0.45 | |||||||||||||||||||||||||||
| He | 0.76 | 0.88 | 0.47 | 0.11 | 0.40 | 0.22 | 0.55 | 0.92 | 0.54 | 0.78 | 0.66 | 0.57 | |||||||||||||||||||||||||||
| FIS | 0.279 | 0.365 | 0.059 | 0.000 | 0.636 | 0.000 | 0.190 | 0.267 | 0.590 | -0.309 | -0.191 | 0.171 | |||||||||||||||||||||||||||
| Phwe | 0.0795 | 0.0317 | 0.5320 | — | 0.1771 | — | 0.1576* | 0.0776* | 0.0133* | 0.1778 | 0.4218 | ||||||||||||||||||||||||||||
| 海 尾 | N | 10 | 7 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 9 | 9 | |||||||||||||||||||||||||||
| RS | 3.7 | 6.0 | 4.4 | 2.9 | 1.0 | 1.0 | 3.6 | 6.1 | 2.8 | 3.0 | 2.0 | 3.3 | |||||||||||||||||||||||||||
| Ho | 0.45 | 0.36 | 0.64 | 0.36 | — | — | 0.18 | 0.36 | 0.00 | 0.73 | 0.73 | 0.42 | |||||||||||||||||||||||||||
| He | 0.52 | 0.86 | 0.63 | 0.54 | — | — | 0.54 | 0.82 | 0.41 | 0.72 | 0.67 | 0.64 | |||||||||||||||||||||||||||
| FIS | 0.138 | 0.333 | -0.007 | 0.333 | — | — | 0.623 | 0.543 | 1.000 | -0.524 | -0.778 | 0.185 | |||||||||||||||||||||||||||
| Phwe | 0.3443 | 0.0665 | 0.2047 | 0.0955 | — | — | 0.0084* | 0.0000* | 0.0022* | 0.2135 | 0.0557 | ||||||||||||||||||||||||||||
| 大 铲 礁 | N | 11 | 11 | 11 | 11 | 11 | 11 | 11 | 11 | 11 | 11 | 11 | |||||||||||||||||||||||||||
| RS | 4.6 | 5.2 | 2.8 | 3.0 | 1.0 | 1.0 | 2.8 | 4.6 | 2.8 | 3.9 | 2.0 | 3.1 | |||||||||||||||||||||||||||
| Ho | 0.18 | 0.45 | 0.45 | 0.82 | — | — | 0.18 | 0.73 | 0.45 | 0.64 | 0.91 | 0.54 | |||||||||||||||||||||||||||
| He | 0.85 | 0.77 | 0.46 | 0.70 | — | — | 0.41 | 0.71 | 0.39 | 0.75 | 0.52 | 0.62 | |||||||||||||||||||||||||||
| FIS | 0.779 | 0.383 | -0.163 | -0.259 | — | — | 0.459 | -0.026 | -0.163 | 0.136 | -0.818 | 0.036 | |||||||||||||||||||||||||||
| Phwe | 0.0000* | 0.0000 | 1.0000 | 0.3486 | — | — | 0.0992 | 0.1568 | 1.0000 | 0.0041 | 0.0196 | ||||||||||||||||||||||||||||
| 龙 湾 | N | 22 | 21 | 21 | 22 | 22 | 22 | 22 | 22 | 21 | 22 | 22 | |||||||||||||||||||||||||||
| RS | 3.2 | 4.8 | 3.5 | 2.3 | 1.3 | 1.0 | 2.9 | 6.2 | 2.0 | 3.6 | 2.0 | 3.0 | |||||||||||||||||||||||||||
| Ho | 0.32 | 0.64 | 0.36 | 0.86 | 0.05 | — | 0.32 | 0.73 | 0.32 | 0.73 | 0.68 | 0.50 | |||||||||||||||||||||||||||
| He | 0.40 | 0.73 | 0.49 | 0.55 | 0.09 | — | 0.54 | 0.85 | 0.38 | 0.70 | 0.51 | 0.52 | |||||||||||||||||||||||||||
| FIS | 0.128 | 0.056 | 0.140 | -0.684 | 0.000 | — | 0.407 | 0.141 | -0.176 | -0.043 | -0.346 | -0.038 | |||||||||||||||||||||||||||
| Phwe | 0.1269 | 0.1715 | 0.0871 | 0.0006 | — | — | 0.0107* | 0.0000 | 1.0000 | 0.0040 | 0.1924 | ||||||||||||||||||||||||||||
| 西 沙 七 连 屿 | N | 13 | 13 | 11 | 13 | 13 | 13 | 13 | 13 | 13 | 12 | 13 | |||||||||||||||||||||||||||
| RS | 1.5 | 5.2 | 3.9 | 1.0 | 1.0 | 1.0 | 2.5 | 8.0 | 2.7 | 3.4 | 2.5 | 3.0 | |||||||||||||||||||||||||||
| Ho | 0.08 | 0.54 | 0.54 | — | — | — | 0.15 | 0.77 | 0.23 | 0.69 | 0.46 | 0.43 | |||||||||||||||||||||||||||
| He | 0.15 | 0.78 | 0.83 | — | — | — | 0.28 | 0.88 | 0.40 | 0.67 | 0.52 | 0.56 | |||||||||||||||||||||||||||
| FIS | 0.000 | 0.320 | 0.146 | — | — | — | 0.461 | 0.127 | 0.333 | -0.356 | 0.007 | 0.130 | |||||||||||||||||||||||||||
| Phwe | — | 0.0070 | 0.0340 | — | — | — | 0.2402 | 0.1348 | 0.0271 | 0.5811 | 1.0000 | ||||||||||||||||||||||||||||
注: N: 个体数; RS: 每个位点的等位基因丰度; Ho: 观察杂合度; He: 期望杂合度; FIS: 近交系数; Phwe: 哈迪温伯格平衡; 加粗表示经 Bonferroni 校正后显著偏离哈迪温伯格平衡(初始 a = 0.05/11 = 0.0045); *表示经 MICRO-CHEKER 检测后显示该位点在群体中存在无效等位基因, 因而可能导致偏离哈迪温伯格平衡; 空白表示无法计算; —表示不可计算 |
表4 澄黄滨珊瑚11个群体遗传多样性参数Tab. 4 Genetic diversity for 11 populations of Porites lutea |
| 群体 | N | RS | Ho | He | FIS | HWE (偏离位点) |
|---|---|---|---|---|---|---|
| 八所 | 15 | 2.8 ± 1.3 | 0.51± 0.25 | 0.56 ± 0.18 | -0.041 | - |
| 雷公岛 | 16 | 3.3 ± 1.8 | 0.35 ± 0.27 | 0.50 ± 0.27 | 0.251 | plo78, pasE062 |
| 木栏头 | 16 | 3.2 ± 1.5 | 0.36 ± 0.27 | 0.52 ± 0.24 | 0.163 | - |
| 铜鼓岭 | 11 | 3.0 ± 1.1 | 0.31 ± 0.33 | 0.59 ± 0.20 | 0.399 | plo37, plo78, pasE056, pasE060, pasE073 |
| 大洲岛 | 10 | 3.4 ± 1.9 | 0.39 ± 0.23 | 0.52 ± 0.26 | 0.022 | - |
| 鹿回头 | 8 | 3.2 ± 1.6 | 0.39 ± 0.22 | 0.57 ± 0.22 | 0.129 | - |
| 海尾 | 10 | 3.3 ± 1.7 | 0.42 ± 0.25 | 0.64 ± 0.15 | 0.185 | pasE056, pasE062 |
| 大铲礁 | 11 | 3.1 ± 1.4 | 0.54 ± 0.26 | 0.62 ± 0.17 | 0.036 | plo7, plo37, pasE073 |
| 邻昌礁 | 9 | 3.7 ± 1.7 | 0.45 ± 0.30 | 0.57 ± 0.26 | 0.171 | - |
| 龙湾 | 22 | 3.0 ± 1.5 | 0.50 ± 0.26 | 0.52 ± 0.21 | -0.038 | plo78, pasE060, pasE073 |
| 西沙七连屿 | 13 | 3.0 ± 2.1 | 0.43 ± 0.25 | 0.56 ± 0.27 | 0.130 | - |
注: N: 个体数; RS: 等位基因丰度; Ho: 观察杂合度; He: 期望杂合度; FIS: 近交系数; HWE: 偏离位点, 即经 Bonferroni 校正后仍偏离 HWE的位点(初始 a = 0.05/11 = 0.0045); -表示无对应位点 |
表5 澄黄滨珊瑚群体间遗传分化指数FST (对角线以下)以及Nei’s遗传距离(对角线以上)Tab. 5 Pair FST (below diagonal) and Nei’genetic distance (above diagonal) between P. lutea populations |
| 八所 | 雷公岛 | 木栏头 | 铜鼓岭 | 大洲岛 | 鹿回头 | 海尾 | 大铲礁 | 邻昌礁 | 龙湾 | 西沙 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 八所 | - | 0.116 | 0.081 | 0.194 | 0.103 | 0.114 | 0.205 | 0.202 | 0.156 | 0.110 | 0.407 |
| 雷公岛 | 0.072 | - | 0.068 | 0.221 | 0.098 | 0.098 | 0.172 | 0.179 | 0.167 | 0.158 | 0.413 |
| 木栏头 | 0.046 | 0.007 | - | 0.191 | 0.089 | 0.083 | 0.201 | 0.193 | 0.140 | 0.126 | 0.393 |
| 铜鼓岭 | 0.105 | 0.091 | 0.090 | - | 0.174 | 0.199 | 0.253 | 0.286 | 0.231 | 0.222 | 0.405 |
| 大洲岛 | 0.037 | 0.018 | 0.024 | 0.075 | - | 0.128 | 0.190 | 0.207 | 0.165 | 0.194 | 0.387 |
| 鹿回头 | 0.042 | 0.017 | 0.016 | 0.091 | 0.021 | - | 0.236 | 0.227 | 0.217 | 0.169 | 0.432 |
| 海尾 | 0.127 | 0.107 | 0.102 | 0.109 | 0.086 | 0.138 | - | 0.195 | 0.144 | 0.184 | 0.395 |
| 大铲礁 | 0.117 | 0.092 | 0.106 | 0.135 | 0.093 | 0.109 | 0.071 | - | 0.178 | 0.193 | 0.403 |
| 邻昌礁 | 0.057 | 0.097 | 0.067 | 0.089 | 0.101 | 0.086 | 0.045 | 0.053 | - | 0.136 | 0.381 |
| 龙湾 | 0.148 | 0.131 | 0.100 | 0.152 | 0.129 | 0.166 | 0.055 | 0.084 | 0.060 | - | 0.408 |
| 西沙 | 0.341 | 0.330 | 0.338 | 0.309 | 0.311 | 0.339 | 0.307 | 0.294 | 0.306 | 0.325 | - |
图3 11个澄黄滨珊瑚群体基于DA (a)和FST*遗传距离(b)的Neighbor-Joining系统树粉色为西岸群体(以及龙湾群体); 蓝色为北东南连通带群体; 数字表示自距值 Fig. 3 Neighbor-joining tree of 11 P. lutea populations based on DA (a) and FST* genetic distance [FST/(1-FST)] (b). The west coast populations and LwR population is shown in red, and the north-south-east connected belt population is shown in blue |
图4 11个澄黄滨珊瑚地理群体基于11个微卫星位点的FST*的主坐标分析蓝圈表示海南岛北东南连通带群体; 红圈表示西岸群体; 十字线为坐标 Fig. 4 Principal coordinate analysis of FST* of 11 microsatellite loci from 11 P. lutea populations. Coordinate 1 mainly reflects the differentiation between XsR population and the populations of Hainan fringing reefs, and coordinate 2 mainly reflects the differentiation between the populations of north-south-east connected belt (blue circle) and the populations of west coast (red circle, including LwR offshore population) |
| [1] |
胡敏航, 陈天然, 张文静, 2018. 滨珊瑚Sr/Ca的种间和种内差异性[J]. 热带海洋学报, 37(6): 74-84.
|
| [2] |
李福宇, 陈丹丹, 李元超, 等, 2021. 澄黄滨珊瑚微卫星标记的开发[J]. 基因组学与应用生物学, 40(7): 2513-2521.
|
| [3] |
刘小菊, 施祺, 陶士臣, 等, 2022. 近165年来中沙环礁中北暗沙滨珊瑚生长率及其对海温变化的响应[J]. 热带海洋学报, 41(5): 64-73.
|
| [4] |
王道儒, 吴瑞, 李元超, 等, 2013. 海南省热带典型海洋生态系统研究[M]. 北京: 海洋出版社:1-210.
|
| [5] |
王建丰, 王毅, 孙双文, 2009. 海南岛西南春季海流特征分析[J]. 中国海洋大学学报(自然科学版), 39(S1): 1-6.
|
| [6] |
张乔民, 2001. 我国热带生物海岸的现状及生态系统的修复与重建[J]. 海洋与湖沼, 32(4): 454-464.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}