
低温胁迫下红榄李(Lumnitzera littorea)DEAD-box RNA解旋酶基因的表达分析
|
郝露露, 女(1996—), 山西省临汾市人, 硕士研究生, 从事红树植物的抗逆研究。email: |
Copy editor: 林强
收稿日期: 2022-03-29
修回日期: 2022-05-23
网络出版日期: 2022-05-31
基金资助
海南省林业科学研究院(海南省红树林研究院)基础性科研工作(KYYS-2021-04)
海南省科研院所技术创新专项(KYYS-2021-13)
海南省科研院所技术创新专项(KYYS-2021-22)
海南省科研院所技术创新专项基础性科研工作项目(jcxk202003)
国家自然科学基金(32071503)
Expression of DEAD-box RNA helicase enzyme genes in Lumnitzera littorea under low temperature stress
Copy editor: LIN Qiang
Received date: 2022-03-29
Revised date: 2022-05-23
Online published: 2022-05-31
Supported by
Basic scientific research work of Hainan Forestry Research Institute (Hainan Mangrove Research Institute)(KYYS-2021-04)
Technological Innovation Special project of Hainan Scientific Research Institute(KYYS-2021-13)
Technological Innovation Special project of Hainan Scientific Research Institute(KYYS-2021-22)
Special Basic Research Work Project for Technological Innovation in Hainan Research Institutes(jcxk202003)
National Natural Science Foundation of China(32071503)
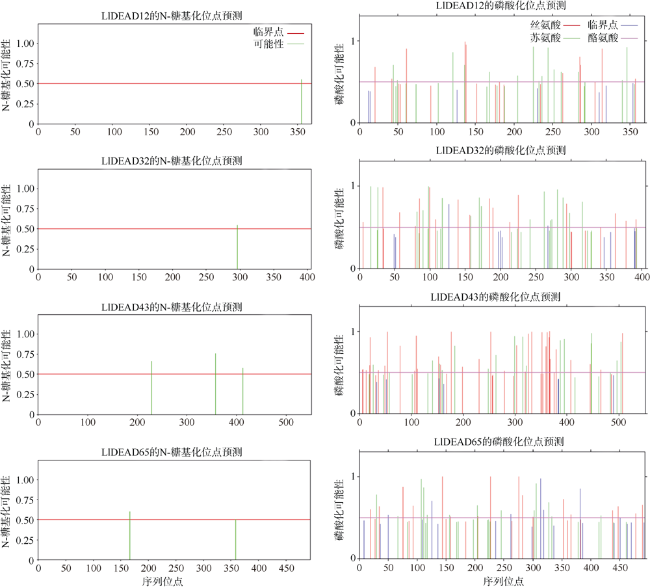
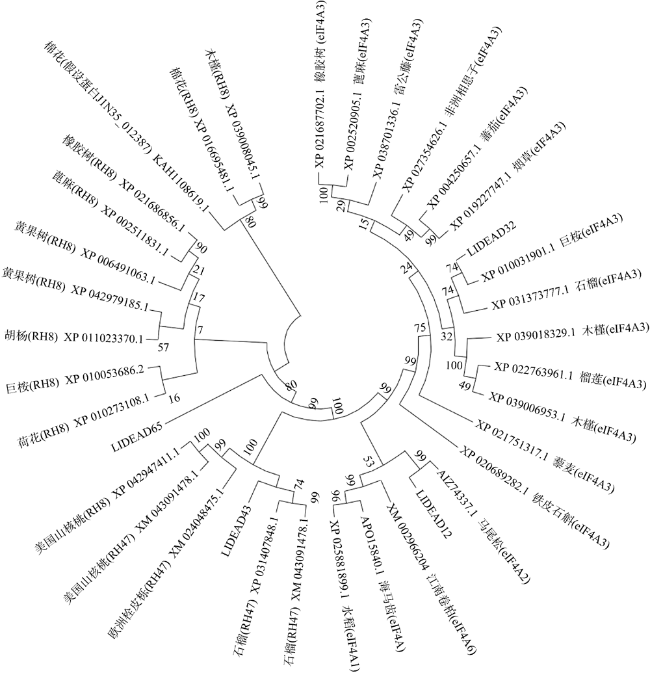
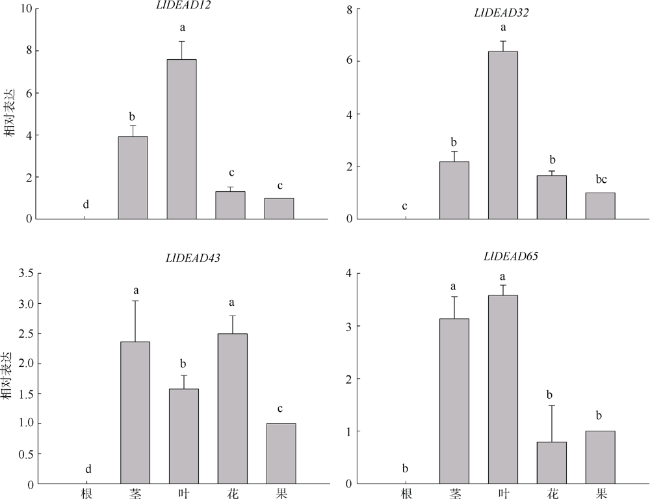
转录组分析发现DEAD-box RNA解旋酶家族参与红榄李(Lumnitzera littorea)对低温胁迫的响应。本文对4个低温显著差异表达基因LlDEAD12, LlDEAD32, LlDEAD43和LlDEAD65的生物信息学特性、组织特异性以及在不同温度处理下的红榄李幼苗中的差异表达进行研究。结果表明, LlDEAD12、LlDEAD32、LlDEAD43和LlDEAD65均属于疏水性蛋白, 二级结构均由α-螺旋、β-转角、延伸链和无规则卷曲组成, 具有多个糖基化位点和磷酸化位点, 且不含有跨膜结构域和信号肽。亚细胞定位分析表明, LlDEAD12和LlDEAD32分别定位于细胞质和细胞核, 而LlDEAD43和LlDEAD65则定位于线粒体上。通过蛋白质氨基酸序列的比对分析发现LlDEAD12、LlDEAD32和LlDEAD43分别与马尾松(Pinus massoniana )、巨桉 (Eucalyptus grandis)和石榴 (Punica granatum)的DEAD-box RNA解旋酶有较近的亲缘关系。荧光定量PCR技术分析发现, LlDEAD12和LlDEAD32在叶中高表达, LlDEAD43和LlDEAD65在茎和花中高表达, 但这4个基因在根中都不表达, 说明这4个基因对红榄李生长发育的调控主要集中在叶、茎和花等器官。低温胁迫对红榄李幼苗这4个基因的表达均有显著的抑制, 说明它们参与了红榄李在低温环境下的分子响应, 其中LlDEAD12和LlDEAD32可能参与了叶绿体的发育, LlDEAD43和LlDEAD65可能参与到了维持线粒体功能的稳定。以上结果可为红榄李抗冷性苗木的培育提供科学依据。
关键词: 红树林; 红榄李; DEAD-box解旋酶; 低温胁迫; 生物信息学分析
郝露露 , 柯明思 , 朱奕秀 , 许燕敏 , 张颖 , 郑春芳 . 低温胁迫下红榄李(Lumnitzera littorea)DEAD-box RNA解旋酶基因的表达分析[J]. 热带海洋学报, 2022 , 41(6) : 44 -55 . DOI: 10.11978/2022061
Transcriptome differential analysis revealed that the DEAD-box RNA helicase family plays an important role in the response of Lumnitzera littorea to low temperature stress. In this study, we analyzed the bioinformatic properties, tissue-specific expression and differential expression of four genes, LlDEAD12, LlDEAD32, LlDEAD43 and LlDEAD65, in L. littorea seedlings under low temperature treatments. The results showed that LlDEAD12, LlDEAD32, LlDEAD43, and LlDEAD65 are hydrophobic proteins with secondary structures consisting of α-helix, β-turn, extended chain and irregular coiling, with many glycosylation sites and phosphorylation sites, and do not contain transmembrane structural domains or signal peptides. Subcellular localization analysis showed that LlDEAD12 and LlDEAD32 are localized in the cytoplasm and nucleus, respectively, while LlDEAD43 and LlDEAD65 are localized in the mitochondria. Comparative analysis of protein amino acid sequences indicated that LlDEAD12, LlDEAD32 and LlDEAD43 had similar affinities to DEAD-box RNA helicase enzymes of Pinus massoniana, Eucalyptus grandis and Punica granatum, respectively. Analysis of the tissue expression specificity of the four genes by fluorescence quantitative PCR revealed that LlDEAD12 and LlDEAD32 highly expressed in leaves, while LlDEAD43 and LlDEAD65 highly expressed in stems and flowers, and none of the four genes expressed in roots, indicating that the regulation of growth and development of L. littorea by these four genes was mainly concentrated in leaf, stem and flower organs. The expression of these four genes was significantly suppressed in L. littorea seedlings under low temperature stress, indicating that these four genes are involved in the molecular response of L. littorea under cold temperature environment, among which LlDEAD12 and LlDEAD32 may be involved in chloroplast development, and LlDEAD43 and LlDEAD65 may be involved in maintaining the stability of mitochondrial function. Our results provide a scientific foundation for the breeding of cold-resistant seedlings of L. littorea.
表1 在线分析的网站及网址Tab.1 Functional website for online analysis and website link |
| 网站 | 功能 | 网址链接 | 参数设置 |
|---|---|---|---|
| ExPASy ProtParam | 理化性质分析 | https://web.expasy.org/protparam/ | 默认参数 |
| PSORT | 亚细胞定位分析 | https://www.genscript.com/psort.html?src=leftbar | 默认参数 |
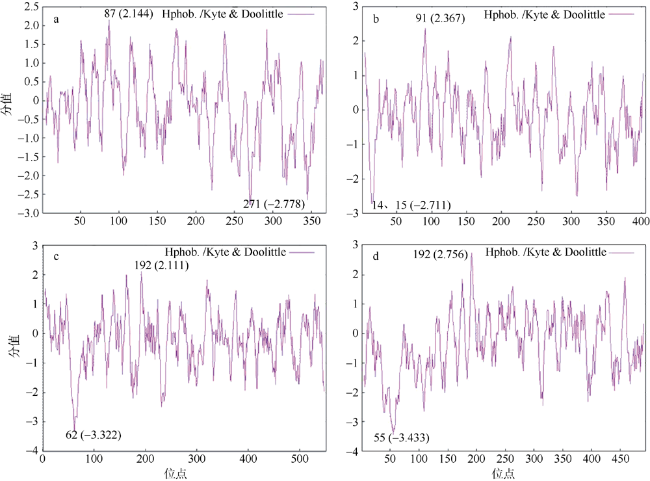
| ProtScale | 亲水性分析 | https://web.expasy.org/cgi-bin/protscale/protscale.pl | Hphob./Kyte & Doolittle, Window size: 9; Weight variation model:linear |
| SOPMA | 二级结构分析 | https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html | Number of conformational states: 4; Similarity threshold: 8; Window width: 17 |
| TMHMM | 跨膜结构分析 | https://services.healthtech.dtu.dk/service.php?TMHMM-2.0 | 默认参数 |
| SWISS-MODEL | 三级结构分析 | https://swissmodel.expasy.org/interactive | 默认参数 |
| SignalP4.1 | 信号肽分析 | https://services.healthtech.dtu.dk/service.php?SignalP-4.1 | Organism group: Eukaryotes; D-cutoff values: Default; Output format: Standard; Method: Input sequences may include TM regions |
| NetPhos3.1 | 磷酸化位点分析 | https://services.healthtech.dtu.dk/service.php?NetPhos-3.1 | Residues to predict: all three; |
| NetNGlyc1.0 | 糖基化位点分析 | https://services.healthtech.dtu.dk/service.php?NetNGlyc-1.0 | 默认设置 |
表2 qPCR引物序列Tab. 2 Primer sequence of qPCR |
| 引物 | 引物序列(5′-3′) | 引物长度/bp |
|---|---|---|
| AthU6-F | ACATCCGATAAAATTGGAACGA | 168 |
| AthU6-R | TTTTTTTGGACCATTTCTCGAT | |
| LlDEAD12-F | GGCTATGACTACCGCAATCTC | 237 |
| LlDEAD12-R | GCCCTTTCAACACTATGCTTCT | |
| LlDEAD32-F | CTGTTGTGCTCTGCCCCA | 204 |
| LlDEAD32-R | CCATGTTCCCATCTTCAATATG | |
| LlDEAD43-F | ATCCTTCCATAGGCGTTCA | 105 |
| LlDEAD43-R | TTCCAGGGGTAGCCACAA | |
| LlDEAD65-F | CGGGCAGATTGGTGGATT | 165 |
| LlDEAD65-R | TGGTCTGTCTTTCACCTCGG |
表3 qPCR反应体系Tab. 3 qPCR reaction system |
| 扩增程序 | 成份 | 体积/μL | |
|---|---|---|---|
| 95℃ 10min | 2xqPCRmix | 10 | |
| 95℃ 10s | 40cycle | F引物(10pmol·μL-1) | 0.5 |
| 60℃ 60s | R引物(10pmol·μL-1) | 0.5 | |
| 95℃ 5s | DNA 模板 | 2 | |
| 60℃ 5s | 每升温0.5℃检测一次 | ddH2O | 7 |
| 95℃ 5s | total | 20 |
表4 红榄李DEAD-box蛋白理化性质分析及亚细胞定位分析Tab. 4 Physicochemical property and subcellular localization |
| 理化指标 | 蛋白质 | |||
|---|---|---|---|---|
| LlDEAD12 | LlDEAD32 | LlDEAD43 | LlDEAD65 | |
| 氨基酸的数量 | 1546 | 2619 | 2144 | 3302 |
| 分子量/Da | 129345.23 | 220714.18 | 176455.64 | 276389.07 |
| 等电点 | 4.94 | 4.82 | 4.94 | 4.82 |
| 稳定指数 | 56.86 | 51.35 | 38.34 | 47.32 |
| 脂肪指数 | 27.04 | 31.54 | 29.24 | 31.8 |
| 亲水性总平均值 | 0.915 | 0.882 | 0.711 | 0.864 |
| 原子总数 | 16283 | 28285 | 22738 | 35465 |
| 亚细胞定位 | 细胞质 | 细胞核 | 线粒体 | 线粒体 |
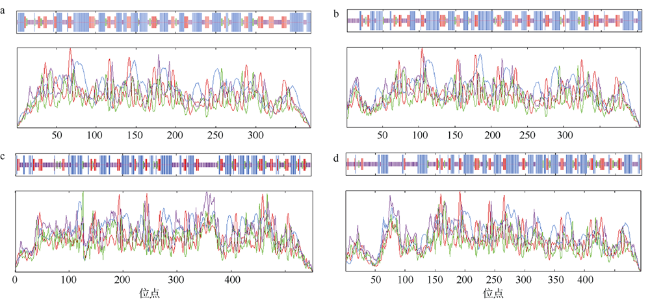
图2 DEAD-box蛋白质的二级结构预测a. LlDEAD12; b. LlDEAD32; c. LlDEA43; d. LlDEAD65。图中蓝色的方块和线条代表α-螺旋; 绿色的方块和线条代表β-折叠; 黄色的方块和线条代表无规则卷曲; 红色的方块和线条代表延伸链 Fig. 2 Predicted secondary structure of DEAD-box protein. (a) LlDEAD12; (b) LlDEAD32; (c) LlDEA43; (d) LlDEAD65. Blue squares and lines in the figure represent α-helices; green squares and lines represent β-sheets; yellow squares and lines represent random coils; red squares and lines represent extended chains |
表5 DEAD-box蛋白质的二级结构类型及占比Tab. 5 Secondary structure types and percentages of DEAD-box proteins |
| α-螺旋 | 延伸链 | β-转角 | 无规则卷曲 | |
|---|---|---|---|---|
| LlDEAD12 | 42.82% | 21.68% | 5.96% | 42.82% |
| LlDEAD32 | 41.38% | 20.69% | 4.68% | 33.25% |
| LlDEAD43 | 33.21% | 16.15% | 6.35% | 44.28% |
| LlDEAD65 | 37.25% | 16.40% | 5.67% | 40.69% |
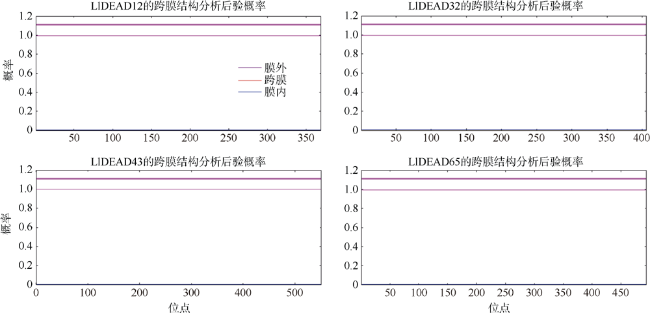
图4 DEAD-box蛋白的跨膜结构预测蓝色线条表示该氨基酸序列在膜内的可能性为0, 在膜外的可能性为100% Fig. 4 Predicted transmembrane structure of DEAD-box protein. The blue line in the figure indicates that the probability of the amino acid sequence in the membrane is 0, and the probability of being outside the membrane is 100%. |
图7 DEAD-box蛋白质的进化关系分析此分子进化树是利用MEGA X进行构建的。分支上的数值表示可信度为1000次重复节点的Bootstrap Fig. 7 Evolutionary relationship analysis. This molecular phylogenetic tree was constructed using MEGA X. The numbers on the branches represent the reliability percent of bootstrap values based on 1000 replication |
图8 DEAD-box基因在不同组织的相对表达水平不同字母表示具有显著性差异(P<0.05) Fig. 8 Relative expression levels of DEAD-box genes in different tissues. a, b, c indicate significant differences in P<0.05 |
图9 DEAD-box基因在不同温度下的相对表达水平第1处理温度为8/5℃; 第2组处理温度为15/12℃; 第3组处理温度为25/23℃; 第4组处理温度为34/30℃。不同字母表示具有显著性差异(P<0.05) Fig. 9 Relative expression levels of DEAD-box genes at different temperatures. Group 1 treated at 8/5°C; Group 2 treated at 15/12°C; Group 3 treated at 25/23°C; Group 4 treated at 34/30°C. a, b, c indicate significant difference in P<0.05 |
| [1] |
蔡敬, 孟小庆, 董婷婷, 等, 2017. DEAD-box解旋酶在植物非生物胁迫响应中的功能研究进展[J]. 生命科学, 29(5): 427-433.
|
| [2] |
陈丹, 彭雄波, 2016. DEAD-box基因家族在拟南芥生长发育中的作用[J]. 植物科学学报, 34(6): 941-948.
|
| [3] |
孔荣荣, 2019. 水稻DEAD-box RNA解旋酶蛋白基因TCD33功能研究[D]. 上海: 上海师范大学.
|
| [4] |
铁原毓, 田洁, 2021. 大蒜蔗糖转化酶基因AsINV的克隆及其响应低温和干旱胁迫的表达分析[J]. 植物生理学报, 57(12): 2258-2270.
|
| [5] |
张颖, 陈光程, 钟才荣, 2021. 中国濒危红树植物研究与恢复现状[J]. 应用海洋学学报, 40(1): 142-153.
|
| [6] |
张颖, 钟才荣, 杨勇, 等, 2018. 濒危红树植物红榄李种质资源挽救[J]. 分子植物育种, 16(12): 4112-4118.
|
| [7] |
钟才荣, 李诗川, 管伟, 等, 2011. 中国3种濒危红树植物的分布现状[J]. 生态科学, 30(4): 431-435.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}