
海水酸化与升温对孔石藻(Porolithon cf. onkodes)生长钙化的影响
|
杨芳芳(1987—), 女, 江西省丰城市人, 博士研究生, 从事藻类生理生态学研究。email: |
Copy editor: 姚衍桃
收稿日期: 2022-05-05
修回日期: 2022-06-02
网络出版日期: 2022-06-15
基金资助
国家自然科学基金(42176157)
中国科学院重点部署项目(ZDRW-XH-2021-2-06)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0402)
中国科学院战略性先导科技专项(A类)(XDA13020203)
The effects of ocean acidification and warming on the growth and calcification in coralline alga Porolithon cf. onkodes
Copy editor: YAO Yantao
Received date: 2022-05-05
Revised date: 2022-06-02
Online published: 2022-06-15
Supported by
National Natural Science Foundation of China(42176157)
Key Programs of the Chinese Academy of Sciences(ZDRW-XH-2021-2-06)
Major Project of Talent Team introduction for Guangdong Provincial Laboratory of Southern Marine Science and Engineering (Guangzhou)(GML2019ZD0402)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13020203)
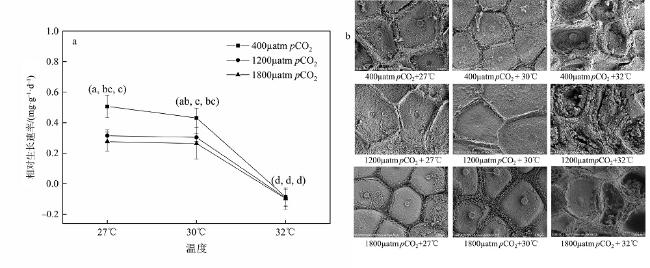
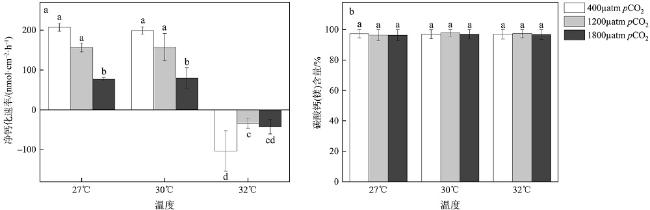
钙化海藻作为一类重要的钙化生物, 不仅能构建稳固礁体, 而且部分藻株能诱导珊瑚幼虫附着变态, 在珊瑚礁生态系统中具有极其重要的生态功能。但是, 面对未来海水酸化与升温的共同作用, 珊瑚藻生理过程会发生怎样的变化, 目前尚不清楚。本文选取了广泛分布于珊瑚礁生态系统中的一种钙化海藻——孔石藻(Porolithon cf. onkodes)为研究对象, 采用3个CO2分压(pCO2: 400µatm、1200µatm、1800µatm)和温度(27℃、30℃、32℃)对其培养1个月, 通过监测藻体生长和钙化等参数的变化情况, 探究海水酸化与升温对活体与死亡的孔石藻的影响。研究结果表明, 当温度从27℃升高到30℃时, 活体孔石藻生长钙化没有受到显著影响; 但当温度提高到32℃时, 藻体生长钙化显著下降, 甚至出现溶解现象, 净钙化速率从206.99nmol·cm-2·h−1 (pCO2: 400µatm; 温度: 27℃)降至-42.22nmol·cm-2·h−1 (pCO2: 1200µatm; 温度 32℃)。类似地, pCO2增加也会显著抑制活体孔石藻的生长钙化, 而且该抑制作用随着CO2分压增加而增强。此外, 升温与酸化对藻体叶绿素a含量和净钙化速率均具有交互作用。与活体孔石藻相比, 死亡的藻体骨骼更易受到高温与酸化影响, 当温度升高至30℃或pCO2提高至1200µatm时, 死亡藻体骨骼净溶解速率均显著增加, 而且高温与酸化的交互作用加速了骨骼溶解速率。研究结果表明了酸化与升温不仅会影响活体孔石藻的生长钙化速率, 而且会加速礁体主要组分(藻体碳酸钙骨骼)的溶解速率, 进而影响到珊瑚礁生态系统。本文研究结果对预测未来气候变化对珊瑚礁生态系统的影响、制定珊瑚礁生态系统保护与修复策略均具有重要的理论价值和生态意义。
杨芳芳 , 肖志梁 , 韦章良 , 黄怡 , 龙丽娟 . 海水酸化与升温对孔石藻(Porolithon cf. onkodes)生长钙化的影响[J]. 热带海洋学报, 2023 , 42(1) : 87 -97 . DOI: 10.11978/2022099
Calcified macroalgae, an important calcifying group widely distributed in reef ecosystems, plays vital roles in the primary productivity and reef frameworks construction. Furthermore, several species of calcified algae can induce the coral larval settlement, which is closely related to the sustained development of coral reef ecosystem. However, it is not clear how calcified algae will response to ocean acidification and warming. In this study, healthy and bleached Porolithon cf. onkodes were exposed to different pCO2 (400 µatm, 1200 µatm, 1800 µatm) and temperatures (27 ℃, 30 ℃, 32 ℃) for one month, respectively. The results showed that the growth rate and net calcification rate of living P. onkodes were not significantly affected when the temperature increased from 27 ℃ to 30 ℃, however, these physiological parameters declined significantly at 32 ℃. The net calcification rate decreased from 206.99 nmol·cm-2·h−1 (400 µatm pCO2 + 27 ℃) to -42.22 nmol·cm-2·h−1 (1200 µatm pCO2 + 32 ℃). Similarly, the enhancement of pCO2 also significantly inhibited the growth and calcification in living P. onkodes. Additionally, warming and acidification had interactive effects on the chlorophyll a content and net calcification rate. Compared with living P. onkodes, dead or bleached algal skeletons were more susceptible to warming and acidification. When the temperature was 30 ℃ or the pCO2 was 1200 µatm, the net dissolution rate of dead algal skeletons increased significantly. Furthermore, the adverse effects of warming were exacerbated when high temperatures coincided with acidification. The results revealed that acidification and warming not only affected the growth and calcification rates of living algae, but also accelerated the dissolution rate of calcium carbonate skeleton, which can affect the coral reef ecosystem. The study may contribute to predict the impacts of climate change on reef ecosystem to protect reef ecosystems.
图1 在不同pCO2和温度条件下培养活体孔石藻(a、b)和死亡孔石藻(c、d)时海水的pH值变化图a Changes of seawater pH values at 8:00 (a, c) and 20:00 (b, d) when Porolithon onkodes were exposed to pCO2 and temperature treatments. (a) and (b) correspond to healthy alga, while (c) and (d) correspond to bleached alga |
表1 升温与酸化对孔石藻生长和色素含量的影响的双因素方差分析结果Tab. 1 Two-way ANOVA of effects of warming and acidification on the relative growth rate and pigment contents of P. onkodes |
| 指标 | 自由度 | 均方 | F值 | P值 (Tukey HSD) | |
|---|---|---|---|---|---|
| 相对生长速率 | Tem | 2 | 0.58 | 147.46 | <0.001 (P27℃&30℃=0.56; P27℃&32℃<0.001; P30℃&32℃<0.001) |
| pCO2 | 2 | 0.05 | 11.40 | <0.001(PL&M=0.01; PL&H<0.001; PM&H=0.63) | |
| Tem & pCO2 | 4 | 0.01 | 2.64 | 0.07 | |
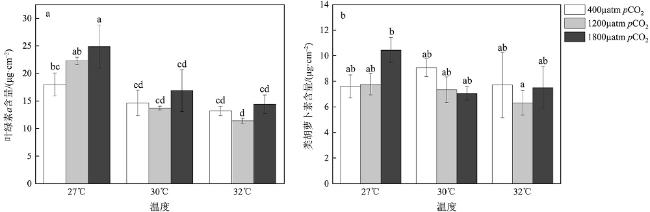
| 叶绿素a含量 | Tem | 2 | 197.83 | 59.12 | <0.001(P27℃&30℃<0.001; P27℃&32℃<0.001; P30℃&32℃=0.21) |
| pCO2 | 2 | 19.59 | 5.85 | <0.001(PL&M=0.82; PL&H=0.01; PM&H=0.04) | |
| Tem & pCO2 | 4 | 12.17 | 3.64 | 0.03 | |
| 类胡萝卜素含量 | Tem | 2 | 4.60 | 2.86 | 0.08 |
| pCO2 | 2 | 3.65 | 2.27 | 0.13 | |
| Tem & pCO2 | 4 | 4.62 | 2.88 | 0.05 | |
注: Tem代表温度, L、M、H分别代表pCO2为400µatm、1200µatm和1800µatm |
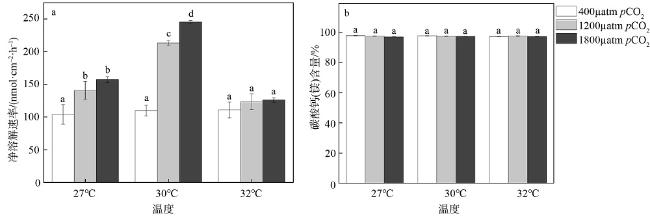
图4 海水酸化与升温对孔石藻净钙化速率(a)和碳酸钙(镁)含量(b)的影响柱子上方小写字母完全不同的表示差异显著(P<0.05) Fig. 4 Effects of acidification and warming on net calcification rate (a), and (Ca, Mg) CO3 content (b) of P. onkodes |
表2 升温与酸化对孔石藻净钙化速率和净溶解速率的影响的双因素方差分析Tab. 2 Two-way ANOVA of effects of warming and acidification on the net calcification rate and dissolution rate of P. onkodes |
| 指标 | 自由度 | 均方 | F值 | P值 (Tukey HSD) | |
|---|---|---|---|---|---|
| 净钙化速率 | Tem | 2 | 126762.19 | 218.46 | <0.001(P27℃&30℃=0.99; P27℃&32℃<0.001; P30℃& 32℃<0.001) |
| pCO2 | 2 | 10572.41 | 18.22 | <0.001(PL&M=0.79; PL&H<0.001; PM&H<0.001) | |
| Tem & pCO2 | 4 | 8770.87 | 15.12 | <0.001 | |
| 净溶解速率 | Tem | 2 | 12152.63 | 133.72 | <0.001(P27℃&30℃<0.001; P27℃&32℃=0.02; P30℃&32℃<0.001) |
| pCO2 | 2 | 11280.92 | 124.13 | <0.001(PL&M<0.001; PL&H<0.001; PM&H=0.003) | |
| Tem & pCO2 | 4 | 3102.12 | 34.13 | <0.001 | |
注: Tem代表温度, L、M、H分别代表pCO2为400µatm、1200µatm和1800µatm |
| [1] |
雷新明, 黄晖, 黄良民, 2012. 珊瑚礁生态系统中珊瑚藻的生态作用研究进展[J]. 生态科学, 31 (5): 585-590.
|
| [2] |
雷新明, 黄晖, 练健生, 等, 2019. 中国珊瑚藻的多样性及分布研究现状[J]. 热带海洋学报, 38(4): 30-40.
|
| [3] |
李银强, 余克服, 王英辉, 等, 2016. 珊瑚藻在珊瑚礁发育过程中的作用[J]. 热带地理, 36(1): 19-26.
|
| [4] |
聂磊, 谢子强, 彭丹, 2021. 海水酸化对珊瑚藻生长和钙化作用的影响[J]. 广东海洋大学学报, 41(3): 67-73.
|
| [5] |
徐智广, 李美真, 霍传林, 等, 2012. 高浓度CO2引起的海水酸化对小珊瑚藻光合作用和钙化作用的影响[J]. 生态学报, 32(3): 699-705.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
IPCC, 2014. Climate change 2013: the physical science basis: working group I contribution to the fifth assessment report of the Intergovernmental Panel on Climate Change[M]. Cambridge: Cambridge University Press.
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}