
红树林固氮微生物及其生态功能研究进展
|
董俊德(1965—), 研究员, 从事微生物海洋学及资源利用研究。email: |
Copy editor: 姚衍桃
收稿日期: 2022-07-20
修回日期: 2022-08-15
网络出版日期: 2022-08-16
基金资助
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0402)
广东省科技计划项目(2020B1212060058)
广东省科技计划项目(2021B1212050023)
中国科学院南海生态环境工程创新研究院创新发展基金项目(ISEE2021ZD03)
Progress on the nitrogen-fixing microorganisms and their ecological functions in mangroves
Copy editor: YAO Yantao
Received date: 2022-07-20
Revised date: 2022-08-15
Online published: 2022-08-16
Supported by
Key Special Project for Exotic Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0402)
Science and Technology Planning Project of Guangdong Province, China(2020B1212060058)
Science and Technology Planning Project of Guangdong Province, China(2021B1212050023)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2021ZD03)
董俊德 , 黄小芳 , 龙爱民 , 王友绍 , 凌娟 , 杨清松 . 红树林固氮微生物及其生态功能研究进展[J]. 热带海洋学报, 2023 , 42(4) : 1 -11 . DOI: 10.11978/2022159
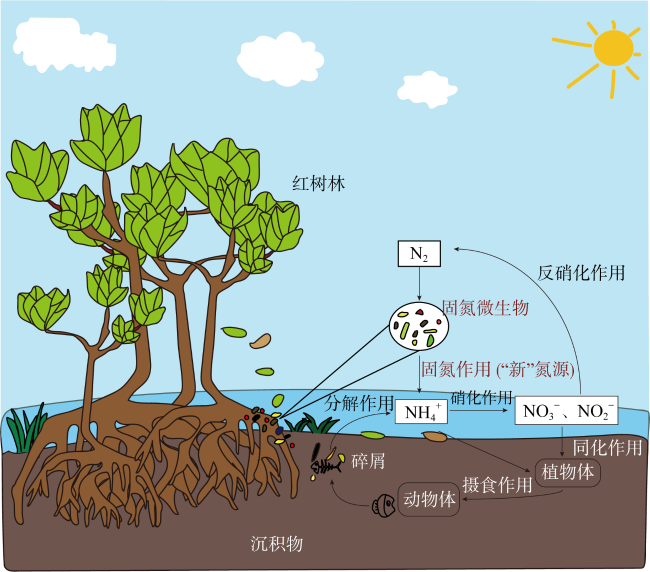
Microbial nitrogen fixation is a crucial part of nitrogen cycling in mangrove ecosystems, it plays an essential role in the nitrogen nutrient supply and primary productivity improvement of ecosystem. This paper reviewed the diazotrophic community structure, nitrogen fixation rate, and the main measurement methods of nitrogen fixation in mangrove ecosystems. We also explored the application of nitrogen-fixing microorganisms in mangrove restoration and the ecological indicators of nitrogen-fixing microorganisms for mangrove habitats, emphasized the vital roles of diazotrophs in coupling carbon, nitrogen, and sulfur cycles in mangrove wetlands, and highlighted new perspectives for nitrogen-fixing microorganisms in the mangrove ecosystem.
表1 2000—2020年红树林生态系统分离的固氮微生物Tab. 1 Nitrogen-fixing microorganisms isolated from mangrove ecosystems from 2000~2020 |
| 序号 | 属名 | 种名 | 分离地点 | 是否为 新属种 | 文献来源 |
|---|---|---|---|---|---|
| 1 | Phyllobacterium | Phyllobacterium sp. | 墨西哥下加利福尼亚州 | 否 | Rojas et al, 2001 |
| 2 | Swaminathania | S. salitolerans | 印度泰米尔纳德邦 | 是 | Loganathan et al, 2004 |
| 3 | Azotobacter | A. chroococcum, A. virelandii, A. beijerinchkii, A. vinelandii,A. chroococcum | 印度帕兰吉佩泰 | 否 | Ravikumar et al, 2004; Kathiresan et al, 2006; Xiong et al, 2016 |
| 4 | Vibrio | V. porteresiae,V. mangrovi,V. plantipsonsor | 印度泰米尔纳德邦 | 是 | Rameshkumar et al, 2008; Rameshkumar et al, 2010b; Rameshkumar et al, 2011 |
| 5 | Teredinibacter | T. turnerae | 巴西塞佩蒂巴湾 | 否 | Trindade-Silva et al, 2009 |
| 6 | Bacillus | B. pumilus | 中国三亚市 | 否 | 凌娟 等, 2010 |
| 7 | Mangrovibacter | M. plantisponsor | 印度泰米尔纳德邦 | 是 | Rameshkumar et al, 2010a |
| 8 | Sporosarcina | S. aquimarina | 印度泰米尔纳德邦 | 否 | Janarthine et al, 2012 |
| 9 | Paenibacillus | Paenibacillus sp. | 中国三亚市 | 否 | Liu et al, 2012 |
| 10 | Stenotrophomonas | Stenotrophomonas sp. | |||
| 11 | Brevundimonas | Brevundimonas sp. | |||
| 12 | Pseudomonas | Pseudomonas spp. | |||
| 13 | Agrobacterium | Agrobacterium sp. | |||
| 14 | Azospirillum | A. lipoferum, A brasilense, A. irakense, A. halopraeferens, A. amazonense | 印度泰米尔纳德邦/中国湛江市 | 否 | Ravikumar et al, 2012; Xiong et al, 2016 |
| 15 | Marinobacterium | M. mangrovicola | 奥地利维也纳 | 是 | Alfaro-Espinoza et al, 2014 |
| 16 | Mangrovibacterium | M. diazotrophicum | 中国三亚市 | 是 | Huang et al, 2014 |
| 17 | Ochrobactrum | O. anthropi | 中国湛江市 | 否 | Xiong et al, 2016 |
| 18 | Synechococcus | S. moorigangaii | 印度孙德尔本斯 | 是 | Singh et al, 2019 |
| 19 | Draconibacterium | D. mangrovi | 中国泉州市 | 是 | Hu et al, 2020 |
| 20 | Maribellus | M. sediminis | 中国泉州市 | 是 | Huang et al, 2020 |
| [1] |
董俊德, 王汉奎, 张偲, 等, 2002. 海洋固氮生物多样性及其对海洋生产力的氮、碳贡献[J]. 生态学报, 22(10): 1741-1749.
|
| [2] |
方安琪, 贺志理, 王成, 等, 2020. 红树林沉积物中微生物驱动硫循环研究进展[J]. 微生物学报, 60(1): 13-25.
|
| [3] |
何雪香, 李玫, 廖宝文, 2012. 红树林固氮菌和解磷菌的分离及对秋茄苗的促生效果[J]. 华南农业大学学报, 33(1): 64-68.
|
| [4] |
凌娟, 董俊德, 张燕英, 等, 2010. 一株红树林根际固氮菌的分离、鉴定以及固氮活性测定[J]. 热带海洋学报, 29(5): 149-153.
|
| [5] |
王荣丽, 廖宝文, 何雪香, 等, 2015. PGPB菌剂对5种红树小苗的野外接菌效应[J]. 东北林业大学学报, 43(1): 103-106.
|
| [6] |
周海霞, 刘其友, 郑经堂, 2013. 海洋微生物固氮作用研究进展[J]. 化学与生物工程, 30(11): 5-9, 30.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}