
|
周林滨(1985—), 男, 山东省鄄城县人, 博士, 从事海洋生态学研究。email: |
|
*感谢匿名审稿专家提出的宝贵修改意见和建议。 |
Copy editor: 林强
收稿日期: 2022-07-07
修回日期: 2022-08-21
网络出版日期: 2022-09-05
基金资助
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0405)
广东省基础与应用基础研究基金项目(2019A1515011645)
国家留学基金资助(202004910004)
中国科学院南海海洋研究所自主部署项目(SCSIO202204)
广东省科技计划项目(2020B1212060001)
Iron-aluminum hypothesis and the potential of ocean aluminum fertilization as a carbon dioxide removal strategy
Copy editor: LIN Qiang
Received date: 2022-07-07
Revised date: 2022-08-21
Online published: 2022-09-05
Supported by
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0405)
Guangdong Basic and Applied Basic Research Foundation(2019A1515011645)
China Scholarship Council(202004910004)
Development Fund of South China Sea Institute of Oceanology of the Chinese Academy of Sciences(SCSIO202204)
Guangdong Provincial Science and technology plan project(2020B1212060001)
人为活动引起的二氧化碳(CO2)等温室气体排放是驱动全球变暖的主要因素。全球变暖对粮食、水资源、能源、经济安全等领域均产生了严重威胁。减缓全球变暖势在必行, 不仅需要大规模减少CO2等温室气体排放, 还需要大量部署CO2移除(carbon dioxide removal, CDR)技术(又称负排放技术), 主动从空气中移除CO2并长期封存, 尽快使全球CO2净排放减少为零, 达到“碳中和”。海洋占地球表面积的70%, 是最大的活跃碳库, 具有巨大的CO2吸收潜力。基于海洋的CDR是实现碳中和的必要途径, 海洋CDR理论、方法、技术研究已成为热点和前沿领域。目前, 海洋CDR的研究认知仍相对较弱, 具有广阔的发展空间。缓解全球变暖的迫切需求促使海洋碳汇基础理论和海洋CDR研究快速发展, 原创性进展不断出现。本文主要综述铁铝假说的理论基础, 探讨基于“海洋铝施肥”的CDR发展潜力。铁铝假说认为, 铝可增强上层海洋浮游植物固碳, 降低生源碳分解速率, 提高海洋生物泵效率, 增加碳向深海输出、封存, 调控海洋碳汇的形成, 影响大气中CO2的浓度。与铁一样, 铝可能也是影响地球历史时期和现代气候变化的关键因子。通过提高铁的利用效率和向深海碳输出效率, 铝可以弥补人工海洋铁施肥的不足, 赋予海洋铝施肥成为新型的基于自然碳汇的CDR方法和技术的潜力。作为一种CDR方法, 海洋铝施肥尽管具有潜在高效的特点, 但还处于较为初级的概念阶段。本文提出, 应当从上层海洋浮游植物固碳、生源碳向深海输出、碳的长期封存三个方面, 进一步研究铝增强海洋碳汇的作用机制, 完善铁铝假说和海洋铝施肥的理论基础; 并在不同时空尺度上检验海洋铝施肥的CDR效能及其潜在环境影响, 为基于海洋铝施肥的CDR技术开发和应用提供科学基础。
周林滨 , 黄良民 , 谭烨辉 . 铁铝假说与海洋铝施肥增汇潜力展望*[J]. 热带海洋学报, 2023 , 42(3) : 1 -18 . DOI: 10.11978/2022153
Human-induced emissions of greenhouse gases such as carbon dioxide (CO2) are the main drivers of global warming. Global warming poses a serious threat to the security of food, water resources, energy, economy, and other fields. Alleviating global warming is imperative. Not only does it require massive greenhouse gas emissions reduction, but also large-scale deployment of carbon dioxide removal (CDR) or negative emissions techniques to intentionally remove CO2 from the air and sequestrate it for a long period so that to decrease global net CO2 emissions to zero as soon as possible, and achieve "carbon neutrality". The ocean accounts for 70% of the earth's surface area and is the largest active carbon pool. It has a huge potential to absorb CO2. Ocean-based CDR is necessary to achieve carbon neutrality. The research on the theory, method, and technology of ocean CDR has become a hot spot and frontier field. At present, the knowledge of ocean CDR is still relatively limited, and there is a large space for development. The urgent need to mitigate global warming is promoting the rapid development of the basic theory of marine carbon sinks and ocean CDR research, and original progress is emerging. This paper mainly summarizes the theoretical basis of the Iron-Aluminum Hypothesis and discusses the potential of ocean aluminum fertilization as a CDR strategy. The iron-aluminum hypothesis indicates that aluminum can enhance carbon fixation by phytoplankton in the upper ocean, reduce the decomposition rate of biogenic carbon, improve the efficiency of the biological pump, increase carbon export and sequestration to the deep sea, regulate marine carbon sinks, and affect the concentration of CO2 in the atmosphere. Thereby, as well as iron, aluminum may be a key factor in influencing historical and modern climate changes. Aluminum improves the efficiency of iron use and carbon export to the deep ocean, which can make up for the shortage of artificial ocean iron fertilization, and endow ocean aluminum fertilization with the potential to become a new CDR method and technology based on natural carbon sinks. Despite its potential high efficiency, ocean aluminum fertilization as a CDR method is still nascent. We suggest further study on the mechanisms underlying the roles of aluminum in enhancing marine carbon sinks from the three aspects 1) carbon fixation by marine phytoplankton in the upper ocean, 2) biogenic carbon export to the deep ocean, and 3) long-term carbon sequestration, and thus to strengthen the theoretical basis of iron-aluminum hypothesis and ocean aluminum fertilization. We also propose to verify the CDR efficacy of ocean aluminum fertilization and its potential environmental impacts at different temporal and spatial scales. The above two works are expected to provide basic scientific knowledge for the development and application of ocean aluminum fertilization as a CDR strategy.
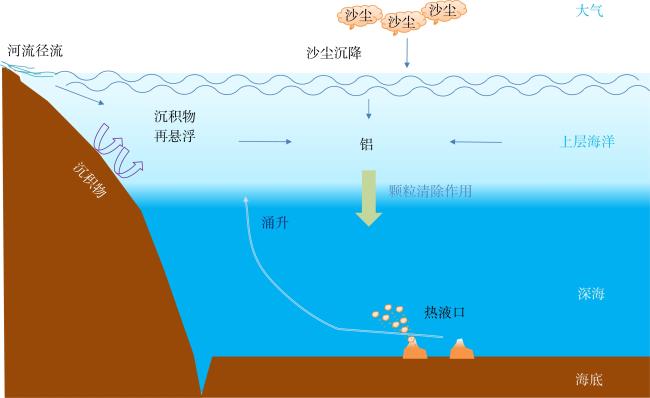
图1 海洋中铝的自然来源示意图改自Zhou等(2018b)。海水中铝的来源主要有河流径流输入, 沙尘气溶胶沉降, 沉积物再悬浮和海底热液口热液喷发。生物或非生物颗粒的吸附、吸收则将铝从海水中清除 Fig. 1 The natural sources of aluminum in the ocean (modified from Zhou et al, 2018b). The main sources of aluminum in the ocean include river runoff, dust deposition, sediment resuspension, and hydrothermal venting. The absorption and/or adsorption by biotic and abiotic particles scavenge aluminum from the seawater |
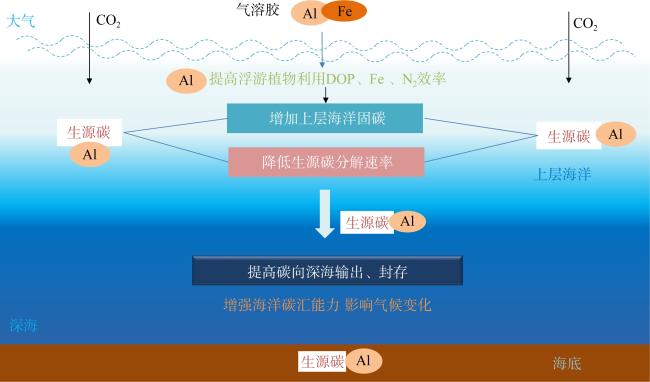
图2 铁铝假说示意图改自Zhou等(2018b)。自然铁施肥过程如(但不限于)沙尘气溶胶沉降不仅向海洋输入铁(Fe)还带来铝(Al)。Al一方面提高海洋浮游植物利用溶解有机磷(dissolved organic phosphorus, DOP)、Fe和N2(固氮)效率, 增加上层海洋固碳; 另一方面降低生源碳分解速率, 提高碳向深海输出、封存, 增强海洋碳汇能力, 从而影响气候变化, 在地球历史时期和现代气候变化过程中发挥重要作用 Fig. 2 Schematic description of the iron-aluminum hypothesis (modified from Zhou et al, 2018b). Natural Fe fertilization processes like (but not limited to) dust deposition provide the ocean not only with Fe but also Al. On the one hand, Al increases carbon fixation by enhancing the utilization efficiency of dissolved organic phosphorus (DOP), iron (Fe), and dinitrogen (N2) by marine phytoplankton. On the other hand, Al can increase carbon export and sequestration by decreasing the decomposition of biogenic carbon. By doing these, Al increases marine carbon sinks and plays an important role in historical (glacial-interglacial) and current climate changes |
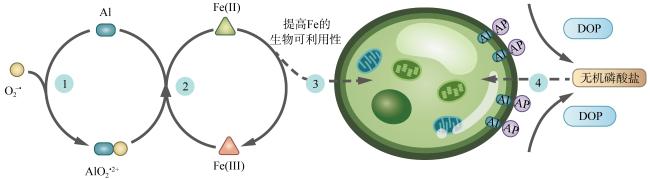
图3 铝(Al)促进海洋浮游植物利用铁(Fe)和溶解有机磷(DOP)营养示意图改自 Zhou等(2021)。过程1: Al与超氧根离子(O2-•)结合形成Al-超氧根络合物(如AlO2•2+); 过程2: Al-超氧根络合物催化三价铁[Fe(Ⅲ)]还原为二价铁[Fe(Ⅱ)]; 过程3: 铁的生物可利用性提高, 增强海洋浮游植物对铁的吸收; 过程4: Al添加使更多的碱性磷酸酶结合在细胞表面, 而不是释放到海水中, 从而有助于细胞在磷限制条件下利用溶解有机磷 Fig. 3 Schematic diagram illustrating how aluminum (Al) may facilitate the uptake of iron (Fe) and the utilization of dissolved organic phosphorus (DOP) by marine phytoplankton. Modified from Zhou et al (2021). 1. Al binds with superoxide (O2-•) to form an Al-superoxide complex (e.g. AlO2•2+); 2. Al-superoxide catalyzes the reduction of ferric iron (Fe(Ⅲ)) to ferrous iron (Fe(Ⅱ)); 3. Increased Fe bioavailability results in enhanced Fe uptake; 4. Al addition leads to a greater association of alkaline phosphatase (AP) with the cell surface rather than being released into seawater and facilitates the cells’ use of DOP in phosphorus-limited conditions |
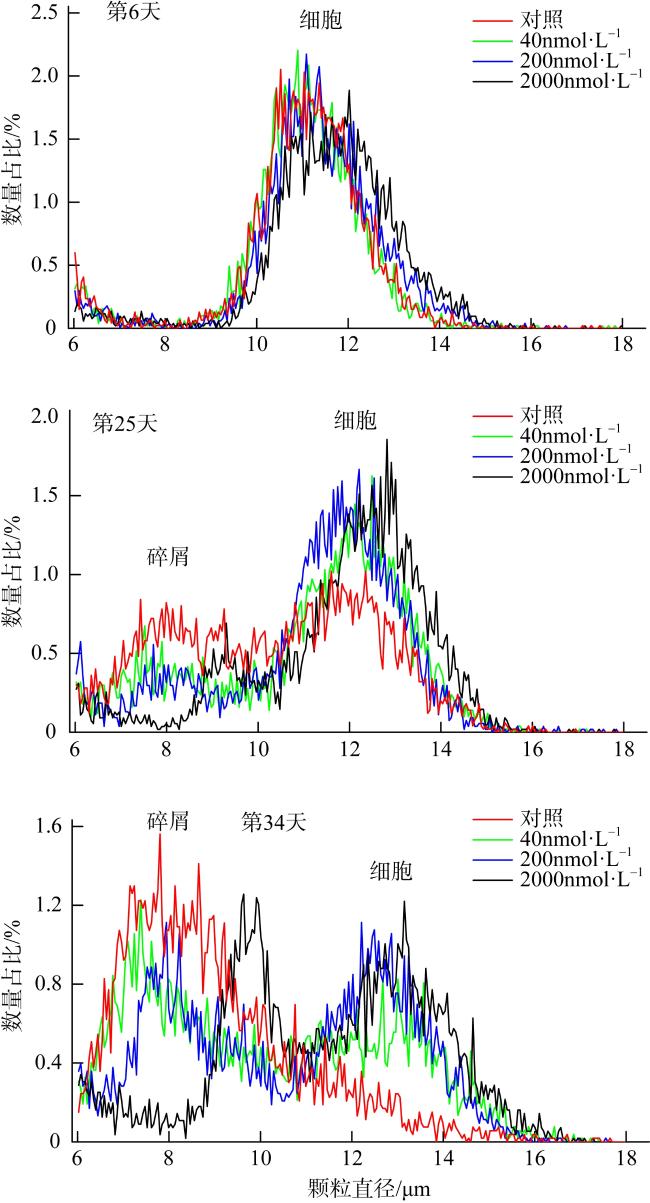
图4 添加不同铝浓度Aquil*培养基中海洋硅藻威氏海链藻细胞颗粒大小分布改自Zhou等(2021)。第6天为指数生长后期, 第25天为衰退期, 第34天为衰退期后期 Fig. 4 Particle size distribution in cultures of the marine diatom Thalassiosira weissflogii in Aquil* media with different aluminum concentrations. Modified from Zhou et al (2021). Day 6, the decline growth phase after the exponential phase; Day 25, the decay phase; Day 34, the late decay phase |
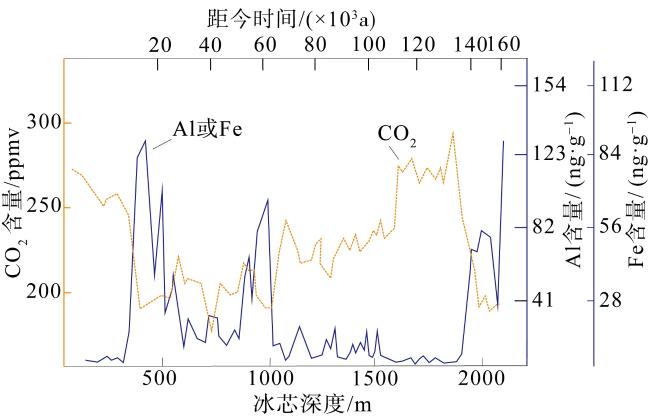
图5 南极Vostok冰芯记录过去16万年铝(Al)、铁(Fe)输入量与冰芯深度、冰芯封存空气CO2浓度变化改自Zhou等(2018b)和Martin (1990)。铁的数据由铝数据根据地壳中两种元素的平均丰度估算(Taylor, 1964) Fig. 5 Antarctic Vostok ice core data show the relationship of aluminum (Al) or iron (Fe) input with ice core depth and mean CO2 concentrations in air tapped into ice in the past 160 thousand years. Modified from Zhou et al (2018b) and Martin (1990). Al data were converted to Fe estimates based on the average crustal abundance values of the two elements (Taylor, 1964) |
| [1] |
焦念志, 2021. 研发海洋“负排放”技术支撑国家“碳中和”需求[J]. 中国科学院院刊, 36(2): 179-187.
|
| [2] |
焦念志, 戴民汉, 翦知湣, 等, 2022. 海洋储碳机制及相关生物地球化学过程研究策略[J]. 科学通报, 67(15): 1600-1606.
|
| [3] |
焦念志, 刘纪化, 石拓, 等, 2021. 实施海洋负排放践行碳中和战略[J]. 中国科学: 地球科学, 51(4): 632-643. (in Chinese)
|
| [4] |
刘纪化, 郑强, 2021. 从海洋碳汇前沿理论到海洋负排放中国方案[J]. 中国科学: 地球科学, 51(4): 644-652. (in Chinese)
|
| [5] |
刘甲星, 周林滨, 柯志新, 等, 2017. 铝对海洋固氮蓝藻 Crocosphaerawatsonii 生长及固氮速率的影响[J]. 热带海洋学报, 36(2): 12-18.
|
| [6] |
骆庭伟, 焦念志, 2022. 海洋负排放:为全球气候治理提供新方案[J]. 前沿科学, 16(2): 83-87 (in Chinese)
|
| [7] |
秦大河, 翟盘茂, 2021, 中国气候与生态环境演变, 2021(第一卷科学基础)[M] // 秦大河, 丁永建, 翟盘茂, 等. 中国气候与生态环境演变评估报告. 北京 (In Chinese).
|
| [8] |
史荣君, 李志红, 周林滨, 等, 2016. 溶解态铝对海洋浮游植物群落结构及聚球藻生长的影响[J]. 南方水产科学, 12(1): 1-8.
|
| [9] |
|
| [10] |
王文涛, 刘纪华, 揭晓蒙, 等, 2022. 海洋支撑碳中和技术体系框架构建的思考与建议[J]. 中国海洋大学学报(自然科学版), 52(3): 1-7.
|
| [11] |
王召伟, 任景玲, 闫丽, 等, 2013. 浮游植物对溶解态Al的清除作用实验研究[J]. 生态学报, 33(22): 7140-7147.
|
| [12] |
杨红生, 丁德文, 2022. 海洋牧场 3. 0: 历程, 现状与展望[J]. 中国科学院院刊, 37(6): 832-839. doi:10.16418/j.issn.1000-3045.20211117011.
|
| [13] |
杨宇峰, 罗洪添, 王庆, 等, 2021. 大型海藻规模栽培是增加海洋碳汇和解决近海环境问题的有效途径[J]. 中国科学院院刊, 36(3): 259-269.
|
| [14] |
于翠平, 潘志强, 陈杰, 等, 2012. 铝对茶树生长与生理特性影响的研究[J]. 植物营养与肥料学报, 18(1): 182-187. (in Chinese)
|
| [15] |
于贵瑞, 郝天象, 朱剑兴, 2022a. 中国碳达峰、碳中和行动方略之探讨[J]. 中国科学院院刊, 37(4): 423-434.
|
| [16] |
于贵瑞, 朱剑兴, 徐丽, 等, 2022b. 中国生态系统碳汇功能提升的技术途径:基于自然解决方案[J]. 中国科学院院刊, 37(4): 490-501.
|
| [17] |
俞志明, 邹景忠, 马锡年, 1995. 粘土矿物去除赤潮生物的动力学研究[J]. 海洋与湖沼, 26(1): 1-6.
|
| [18] |
瞿剑, 2021. 我国首个海上二氧化碳封存示范工程启动[N]. 科技日报, 2021-08-31, pp. 002. (in Chinese)
|
| [19] |
张继红, 刘纪化, 张永雨, 等, 2021. 海水养殖践行 “海洋负排放” 的途径[J]. 中国科学院院刊, 36(3): 252-258.
|
| [20] |
张武昌, 孙松, 2002. 铁假说和HNLC海区的现场铁加富实验[J]. 地球科学进展, 17(4): 613-616.
|
| [21] |
周林滨, 谭烨辉, 黄良民, 2012. 沙尘气溶胶沉降对南海浮游植物生长影响初探[C]. // 中国海洋湖沼学会水环境分会中国环境科学学会海洋环境保护专业委员会, 2012年学术年会论文摘要集. 延吉: 中国海洋湖沼学会. (in Chinese)
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
DE ANDRADE L R M,
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
GESAMP,
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
IOC-R, 2021. Integrated Ocean Carbon Research: A Summary of Ocean Carbon Research, and Vision of Coordinated Ocean Carbon Research and Observations for the Next Decade[R/OL]. [2022-07-07]. https://unesdoc.unesco.org/ark:/48223/pf0000376708
|
| [66] |
IPCC, 2014. 2013 Supplement to the 2006 IPCC Guidelines for National Greenhous Gas Inventories: Wetlands[R/OL]. [2022-07-07]. https://www.ipcc.ch/publication/2013-supplement-to-the-2006-ipcc-guidelines-for-national-greenhouse-gas-inventories-wetlands/
|
| [67] |
IPCC, 2018. Global Warming of 1. 5℃[R/OL]. [2022-07-07]. https://www.ipcc.ch/sr15/
|
| [68] |
IPCC, 2022. Climate change 2022: Mitigation of climate change[R/OL]. [2022-07-07]. https://www.ipcc.ch/report/ar6/wg3/
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
LIU, JIHUA,
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
MENZEL BARRAQUETA J-L,
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
THE ASPEN INSTITUTE ENERGY AND ENVIRONMENTAL PROGRAM, 2021. Guidance for ocean-based carbon dioxide removal projects[R/OL]. (2021-12-08) [2022-07-07]. https://www.aspeninstitute.org/wp-content/uploads/files/content/docs/pubs/120721_Ocean-Based-CO2-Removal_E.pdf
|
| [133] |
THE NATIONAL ACADEMIES OF SCIENCES ENGINEERING AND MEDICINE, 2021. A Research Strategy for Ocean-based Carbon Dioxide Removal and Sequestration[M/OL]. [2022-07-07]. https://doi.org/10.17226/26278
|
| [134] |
|
| [135] |
VAN HULTEN M M P,
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
WORLD HEALTH ORGANIZATION, 2003. Aluminium in drinking-water: background document for development of WHO Guidelines for drinking-water quality. Geneva, Switzerland, World Health Organization.
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}