
海水酸化和升温对两种造礁石珊瑚生长和钙化的影响*
*感谢珊瑚生物学和珊瑚礁生态学学科组全体人员对本论文的完成提供的帮助。
|
周伟华(1976—), 男, 浙江省东阳市人, 博士, 研究员, 从事海洋生态环境研究。email: |
Copy editor: 林强
收稿日期: 2022-07-29
修回日期: 2022-08-30
网络出版日期: 2022-09-07
基金资助
广东省基础与应用基础研究基金(2022A1515010656)
国家自然科学基金(31370500)
海南省重点研发计划项目(ZDYF2020200)
Effects of ocean acidification and warming on the growth and calcification of two reef-building corals*
Copy editor: LIN Qiang
Received date: 2022-07-29
Revised date: 2022-08-30
Online published: 2022-09-07
Supported by
Guangdong Basic and Applied Basic Research Foundation(2022A1515010656)
National Natural Science Foundation of China(31370500)
Key Research and Development Project of Hainan Province(ZDYF2020200)
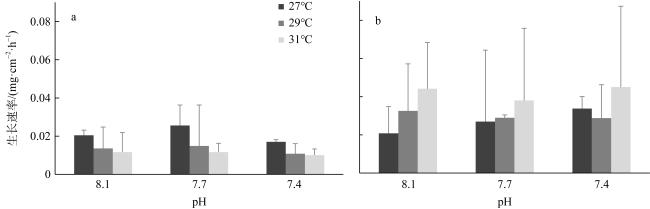
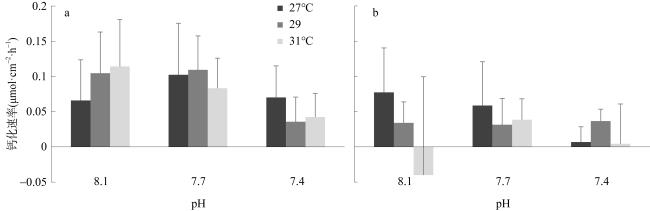
以海南岛三亚鹿回头两种常见的造礁石珊瑚(鹿角杯形珊瑚Pocillopora damicornis 和丛生盔形珊瑚Galaxea fascicularis)为研究对象, 通过室内培养实验探究海水酸化和升温两种联合因子对造礁石珊瑚生长和钙化的影响。结果显示: 两种珊瑚受海水酸化的影响显著, 丛生盔形珊瑚在pH 8.1下的生长速率显著高于pH 7.7和7.4环境, 鹿角杯形珊瑚在pH 7.4下的钙化速率显著低于pH 7.7和8.1环境。且鹿角杯形珊瑚的生长速率与钙化速率显示各温度和 pH 因子之间均出现拮抗现象, 能有效缓解珊瑚白化; 相同的拮抗现象也出现在丛生盔形珊瑚的钙化速率上, 但在其生长速率则表现出协同作用, 加速珊瑚白化。鹿角杯形珊瑚与丛生盔形珊瑚的生长速率和钙化速率对海水升温和酸化的响应存在种间特异性。
周伟华 , 李颖心 , 郭亚娟 , 霍嘉欣 , 宋严 , 朱晴 , 袁翔城 , 刘胜 , 黄晖 . 海水酸化和升温对两种造礁石珊瑚生长和钙化的影响*[J]. 热带海洋学报, 2024 , 43(3) : 49 -57 . DOI: 10.11978/2022171
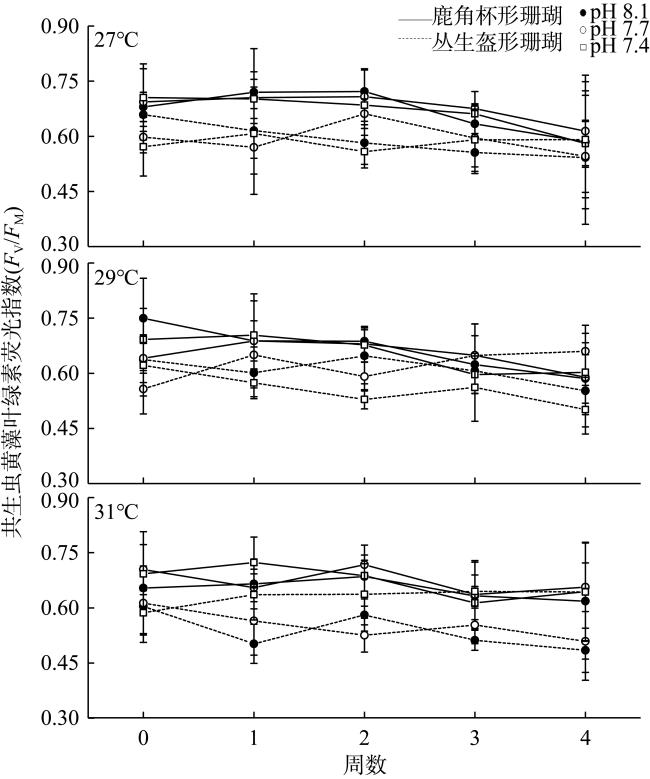
To explore the effects of ocean warming and acidification on coral growth and calcification rate, two widespread coral species, Pocillopora damicornis and Galaxea fascicularis, in the Luhuitou area of the Sanya Bay, were selected for a 28-days controlled incubation experiment. With the interactive treatments of temperatures and pCO2, our results indicated that corals were significantly affected by seawater acidification. Moreover, acidification affected strongly the growth rate of G. fascicularis and the calcification rate of P. damicornis. The chlorophyll fluorescence index (Fv/Fm) of the symbiotic zooxanthellae in P. damicornis was significantly higher than G. fascicularis. Interestingly, the quantitative study showed that there was an antagonistic effect between temperature and pH at some levels, which could effectively alleviate coral bleaching. Thus, we concluded that the growth and calcification rate exhibited interspecific specificity in response to seawater warming and acidification.
Key words: Pocillopora damicornis; Galaxea fascicularis; acidification; warming; growth; calcification
表1 培养实验环境参数列表Tab.1 Environmental parameter values in the incubation experiments |
| pH | 温度/℃ | 盐度/‰ | TA/ (μmol·L-1) | DIC/(μmol·L-1) | pCO2 /μatm | ΩA |
|---|---|---|---|---|---|---|
| 8.1±0.14 | 27±1 | 34±0.02 | 2239±13 | 1888 | 321 | 3.87 |
| 29±1 | 34±0.02 | 2239±13 | 1869 | 318 | 4.12 | |
| 31±1 | 34±0.02 | 2239±13 | 1859 | 317 | 4.25 | |
| 7.7±0.16 | 27±1 | 34±0.02 | 2225±14 | 2091 | 960 | 1.83 |
| 29±1 | 34±0.02 | 2225±14 | 2079 | 962 | 1.97 | |
| 31±1 | 34±0.02 | 2225±14 | 2073 | 963 | 2.04 | |
| 7.4±0.18 | 27±1 | 34±0.02 | 2224±11 | 2197 | 2043 | 0.98 |
| 29±1 | 34±0.02 | 2224±11 | 2189 | 2056 | 1.06 | |
| 31±1 | 34±0.02 | 2224±11 | 2185 | 2062 | 1.10 |
注: TA为总碱度(total alkalinity); DIC为溶解无机碳(dissolved inorganic carbon); ΩA表示CaCO3饱和度, 当ΩA>1 时, 海水中碳酸钙是过饱和的, 有利于钙化沉积作用发生; ΩA=1时, CaCO3的沉淀与溶解达到平衡状态; ΩA<1 则表明海水中碳酸钙为未饱和状态而有溶解的趋势 |
表2 升温处理对酸化与生长速率相互作用的缓解或促进情况Tab. 2 Alleviation or promotion of the interaction of ocean acidification and growth rate by ocean warming treatments |
| 珊瑚物种 | 处理组 | 生长速率 /(mg·cm-2·h-1) | ||
|---|---|---|---|---|
| pH 8.1→7.7(轻度) | pH 7.7→7.4(中度) | pH 8.1→7.4(深度) | ||
| 鹿角杯形珊瑚 | 27℃(处理1) | -0.0051 | 0.0085 | 0.0034 |
| 29℃(处理2) | -0.0013(+) | 0.0040(-) | 0.0028(-) | |
| 31℃(处理3) | 0.0000(++) | 0.0015(- -) | 0.0016(- -) | |
| 丛生盔形珊瑚 | 27℃(处理1) | -0.0061 | -0.0068 | -0.0129 |
| 29℃(处理2) | 0.0036(+) | 0.0002(+) | 0.0038(+) | |
| 31℃(处理3) | 0.0061(++) | -0.0079(- -) | -0.0009(+) | |
注: +表示与处理1比较, 处理2缓解/促进酸化对生长速率的负面/正面影响; ++表示与处理1和处理2相比, 处理3均缓解/促进酸化对生长速率的负面/正面影响; -表示与处理1比较, 处理2加剧/减少酸化对生长速率的负面/正面影响; - -表示与处理1和处理2相比, 处理3均加剧/减少酸化对生长速率的负面/正面影响 |
表3 酸化处理对升温与生长速率相互作用的缓解或促进情况Tab. 3 Alleviation or promotion of the interaction of ocean warming and growth rate by ocean acidification treatments |
| 珊瑚物种 | 处理组 | 生长速率 /(mg·cm-2·h-1) | ||
|---|---|---|---|---|
| 27℃→29℃(轻度) | 29℃→31℃(中度) | 27℃→31℃(深度) | ||
| 鹿角杯形珊瑚 | 8.1(处理1) | 0.0069 | 0.0020 | 0.0088 |
| 7.7(处理2) | 0.0107(+) | 0.0033(+) | 0.0140(+) | |
| 7.4(处理3) | 0.0062(- -) | 0.0007(- -) | 0.0070(- -) | |
| 丛生盔形珊瑚 | 8.1(处理1) | -0.0117 | -0.0116 | -0.0233 |
| 7.7(处理2) | -0.0020(+) | -0.0091(+) | -0.0111(+) | |
| 7.4(处理3) | 0.0050(++) | -0.0163(- -) | -0.0112(+) | |
注: +表示与处理1比较, 处理2缓解/促进升温对生长速率的负面/正面影响; ++表示与处理1和处理2相比, 处理3均缓解/促进升温对生长速率的负面/正面影响; -表示与处理1比较, 处理2加剧/减少升温对生长速率的负面/正面影响; - -表示与处理1和处理2相比, 处理3均加剧/减少升温对生长速率的负面/正面影响 |
表4 升温处理对酸化与钙化速率相互作用的缓解或促进情况Tab. 4 Alleviation or promotion of the interaction of ocean acidification and calcification rate by ocean warming treatments |
| 珊瑚物种 | 处理组 | 钙化速率 /(μmol·cm-2·h-1) | ||
|---|---|---|---|---|
| pH 8.1→7.7(轻度) | pH 7.7→7.4(中度) | pH 8.1→7.4(深度) | ||
| 鹿角杯形珊瑚 | 27℃(处理1) | -0.0366 | 0.0322 | -0.0043 |
| 29℃(处理2) | -0.0049(+) | 0.0737(+) | 0.0688(+) | |
| 31℃(处理3) | 0.0310(++) | 0.0409(+) | 0.0719(++) | |
| 丛生盔形珊瑚 | 27℃(处理1) | 0.0186 | 0.0518 | 0.0704 |
| 29℃(处理2) | 0.0026 (-) | -0.0051(-) | -0.0025(-) | |
| 31℃(处理3) | -0.0785(--) | 0.0340(-) | -0.0445(--) | |
注: +表示与处理1比较, 处理2缓解/促进酸化对钙化速率的负面/正面影响; ++表示与处理1和处理2相比, 处理3均缓解/促进酸化对钙化速率的负面/正面影响; -表示与处理1比较, 处理2加剧/减少酸化对钙化速率的负面/正面影响; - -表示与处理1和处理2相比, 处理3均加剧/减少酸化对钙化速率的负面/正面影响 |
表5 酸化处理对升温与钙化速率相互作用的缓解或促进情况Tab. 5 Alleviation or promotion of the interaction of ocean warming and calcification rate by ocean acidification treatments |
| 珊瑚物种 | 处理组 | 钙化速率 /(μmol·cm-2·h-1) | ||||
|---|---|---|---|---|---|---|
| 27℃→29℃(轻度) | 29℃→31℃(中度) | 27℃→31℃(深度) | ||||
| 鹿角杯形珊瑚 | 8.1(处理1) | -0.0432 | -0.0742 | -0.1173 | ||
| 7.7(处理2) | -0.0272(+) | 0.0069(+) | -0.0202(+) | |||
| 7.4(处理3) | 0.0298(++) | -0.0322(++) | -0.0024(++) | |||
| 丛生盔形珊瑚 | 8.1(处理1) | -0.0386 | -0.0097 | -0.0483 | ||
| 7.7(处理2) | -0.0070(+) | 0.0262(+) | 0.0192(+) | |||
| 7.4(处理3) | 0.0345(++) | -0.0066(+) | 0.0279(++) | |||
注: +表示与处理1比较, 处理2缓解/促进升温对钙化速率的负面/正面影响; ++表示与处理1和处理2相比, 处理3均缓解/促进升温对钙化速率的负面/正面影响; -表示与处理1比较, 处理2加剧/减少升温对钙化速率的负面/正面影响; - -表示与处理1和处理2相比, 处理3均加剧/减少升温对钙化速率的负面/正面影响 |
| [1] |
郭亚娟, 周伟华, 袁翔城, 等, 2018. 两种造礁石珊瑚对海水酸化和溶解有机碳加富的响应[J]. 热带海洋学报, 37(1): 57-63.
|
| [2] |
黄晖, 俞晓磊, 雷新明, 等, 2020. 环境变化对造礁石珊瑚营养方式的影响及其适应性[J]. 海洋科学进展, 38(2): 189-198.
|
| [3] |
江雷, 黄晖, 张浴阳, 等, 2016. 海水升温对壮实鹿角珊瑚幼虫存活和附着的影响[J]. 应用海洋学学报, 35(2): 217-222.
|
| [4] |
李淑, 余克服, 施祺, 等, 2008. 海南岛鹿回头石珊瑚对高温响应行为的实验研究[J]. 热带地理, 28(6): 534-539.
|
| [5] |
刘旭, 黄雯, 俞小鹏, 等, 2022. 适度热胁迫对造礁石珊瑚热耐受性影响的研究[J]. 海洋湖沼通报, 44(1): 99-105.
|
| [6] |
骆雯雯, 梁甲元, 余克服, 等, 2019. 涠洲岛两种石珊瑚在高温胁迫下共生细菌群落结构变化特征[J]. 广西科学, 26(3): 299-307.
|
| [7] |
时翔, 谭烨辉, 黄良民, 等, 2008. 磷酸盐胁迫对造礁石珊瑚共生虫黄藻光合作用的影响[J]. 生态学报, 28(6): 2581-2586.
|
| [8] |
孙有方, 江雷, 雷新明, 等, 2020. 海洋酸化、暖化对两种鹿角珊瑚幼虫附着及幼体存活的影响[J]. 海洋学报, 42(4): 96-103.
|
| [9] |
谭红建, 蔡榕硕, 颜秀花, 2018. 基于CMIP5预估21世纪中国近海海洋环境变化[J]. 应用海洋学学报, 37(2): 151-160.
|
| [10] |
张成龙, 黄晖, 黄良民, 等, 2012. 海洋酸化对珊瑚礁生态系统的影响研究进展[J]. 生态学报, 32(5): 1606-1615.
|
| [11] |
赵美霞, 余克服, 张乔民, 等, 2008. 三亚鹿回头石珊瑚物种多样性的空间分布[J]. 生态学报, 28(4): 1419-1428.
|
| [12] |
郑新庆, 郭富雯, 刘昕明, 等, 2015. 海洋酸化没有显著影响成体鹿角杯形珊瑚的钙化作用和光合能力[J]. 海洋学报, 37(10): 59-68.
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
IPCC, 2014. Climate change 2013: the physical science basis: working Group I contribution to the Fifth assessment report of the Intergovernmental Panel on Climate Change[M]. Cambridge: Cambridge University Press: 255-316.
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
VAN DER ZANDE R M,
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}