
南海热带岛礁生物土壤结皮中细菌的分离及其固砂特性初步研究
|
黄谕(1997—), 女, 海南省文昌市人, 硕士研究生, 从事岛礁生物土壤结皮微生物生态学研究。email: |
Copy editor: 林强
收稿日期: 2022-12-29
修回日期: 2023-02-27
网络出版日期: 2023-03-06
基金资助
国家自然科学基金青年科学基金项目(42206155)
广东省自然科学基金面上项目(2022A1515011889)
广州市科技局基础与应用基础研究项目(202201010499)
Isolation and characterization of sand fixation ability of bacteria in biological soil crusts of the tropical islands, South China Sea
Copy editor: LIN Qiang
Received date: 2022-12-29
Revised date: 2023-02-27
Online published: 2023-03-06
Supported by
National Natural Science Foundation of China(42206155)
Natural Science Foundation of Guangdong Province, China(2022A1515011889)
Guangzhou Basic and Applied Basic Research Foundation(202201010499)
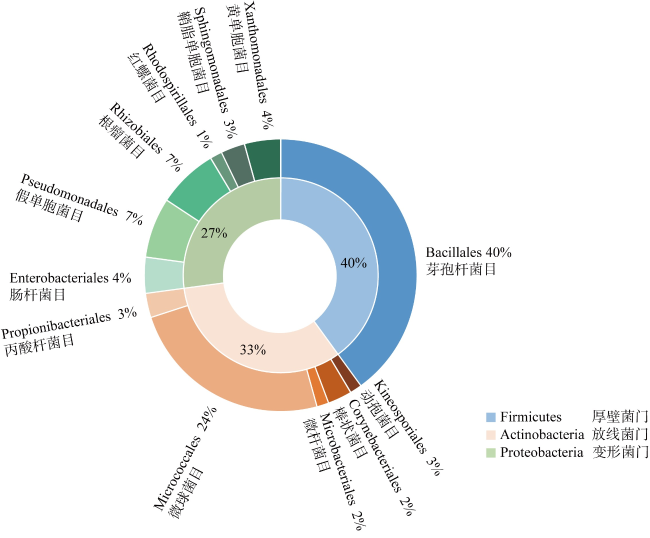
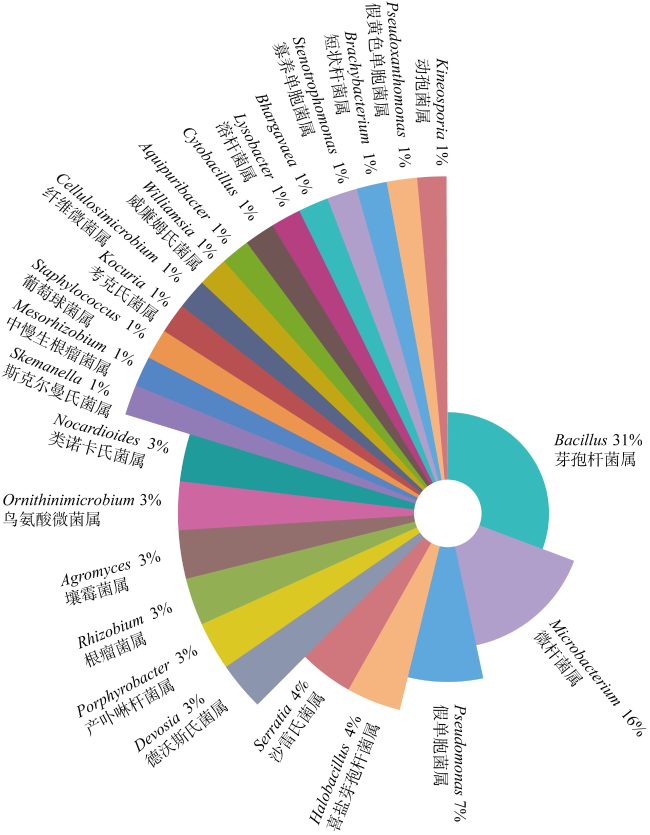
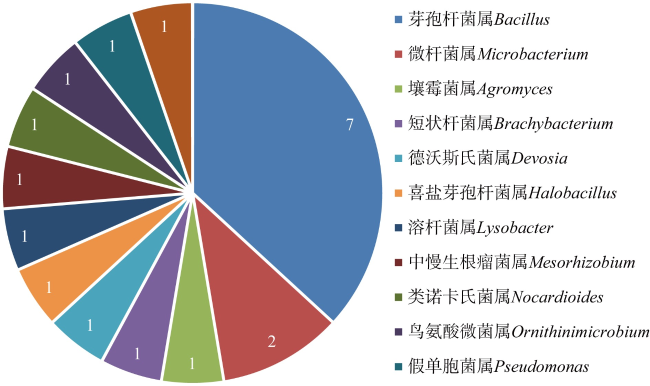
南海热带岛礁生物土壤结皮中有大量可分泌胞外多糖的微生物, 这一特性对固砂具有重要的促进作用。本研究利用TSA、改良TSA、MA三种培养基对采自南海永暑礁及三亚鹿回头海岸带的生物土壤结皮进行可培养细菌的分离纯化。本研究共分离纯化70株细菌, 并进行基于16S rRNA基因序列的鉴定, 发现所分离的菌株属于3门5纲12目19科25属, 厚壁菌门(Firmicutes)为优势门, 芽孢杆菌属(Bacillus)为优势属。此外, 有22株菌与已知物种16S rRNA基因相似性低于98.65%, 为潜在的新物种。纯化菌株的胞外多糖采用乙醇沉淀法提取、苯酚硫酸法检测含量, 发现19株菌胞外多糖含量高于0.013mg·mL-1。选取其中胞外多糖含量最高的9株菌进行土壤团聚能力的测定, 发现将菌株SCSIO 17111(Lysobacter sp.)菌液喷洒于珊瑚砂表面能够使土壤团聚体保持稳定。针对粒径小于0.2mm的细砂, 菌株SCSIO 17111固结的砂土在经过干筛处理后, 保留量可达92.7%。本研究可为岛礁珊瑚砂的固定、减少土壤流失提供高产胞外多糖菌株资源, 还可以为促进岛礁生物土壤结皮的形成提供新的研究思路和方法。
黄谕 , 王琳 , 麦志茂 , 李洁 , 张偲 . 南海热带岛礁生物土壤结皮中细菌的分离及其固砂特性初步研究[J]. 热带海洋学报, 2023 , 42(6) : 101 -110 . DOI: 10.11978/2022266
There are many microbial resources present in biological soil crusts of tropical reefs in the South China Sea, and the extracellular polysaccharides secreted by these microorganisms play an important role in promoting sand consolidation. In this study, three media, TSA, modified TSA and MA, were used to isolate and purify culturable bacteria from biological soil crusts in the Yongshu Reef and Luhuitou, Sanya, South China Sea. A total of 70 bacterial strains were isolated and purified in this study. And based on 16S rRNA gene identification, these isolated strains belonged to 3 phyla, 5 classes, 12 orders and 19 families, 25 genera. The dominant phylum was Firmicutes and the dominant genus was Bacillus. In addition, 22 strains had less than 98.65%16S rRNA gene similarities with known species, which were potential novel species. The extracellular polysaccharide content of purified strains was measured by ethanol precipitation method and phenol sulfate method, and the contents of 19 strains were more than 0.013 mg·mL-1. Finally, 9 strains with the highest extracellular polysaccharide content were selected to determine the soil agglomeration ability. The results showed that strain SCSIO 17111 (Lysobacter) was the only strain which was able to maintain the stability of the biological soil crust clods by spraying. Our study provides high extracellular polysaccharide-producing strain resources for island reef management.
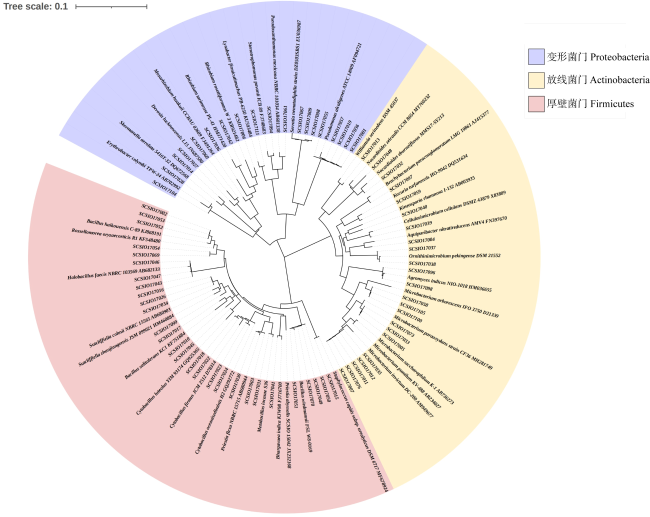
图3 采用邻接法基于16S rRNA基因序列构建的70个菌株系统发育树Fig. 3 Phylogenetic tree of 70 strains constructed by neighbor-joining method based on 16S rRNA gene sequences |
表1 潜在新物种Tab. 1 Potential novel species |
| 菌株编号 | 最近缘物种 | 16S rRNA基因相似性/% |
|---|---|---|
| SCSIO 17100 | Microbacterium arborescens | 98.55 |
| SCSIO 17105 | Microbacterium arborescens | 98.55 |
| SCSIO 17035 | Microbacterium saccharophilum | 98.48 |
| SCSIO 17034 | Bacillus zhanjiangensis | 98.36 |
| SCSIO 17007 | Microbacterium pumilum | 98.33 |
| SCSIO 17026 | Bacillus zhanjiangensis | 98.29 |
| SCSIO 17060 | Devosia lucknowensis | 98.28 |
| SCSIO 17011 | Microbacterium saccharophilum | 98.26 |
| SCSIO 17054 | Bacillus oryzaecorticis | 98.24 |
| SCSIO 17069 | Bacillus oryzaecorticis | 98.23 |
| SCSIO 17076 | Microbacterium invictum | 98.18 |
| SCSIO 17016 | Bacillus zhanjiangensis | 98.15 |
| SCSIO 17014 | Skermanella aerolata | 97.98 |
| SCSIO 17004 | Aquipuribacter nitratireducens | 97.47 |
| SCSIO 17099 | Bacillus cohnii | 97.21 |
| SCSIO 17017 | Bacillus salitolerans | 97.02 |
| SCSIO 17048 | Nocardioides zeicaulis | 96.94 |
| SCSIO 17036 | Mesorhizobium huakuii | 96.93 |
| SCSIO 17031 | Bacillus herbersteinensis | 96.85 |
| SCSIO 17027 | Devosia lucknowensis | 96.66 |
| SCSIO 17010 | Bacillus abyssalis | 96.56 |
| SCSIO 17040 | Kineosporia rhamnosa | 95.90 |
表2 菌株胞外多糖粗测产量Tab. 2 Extracellular polysaccharide productions by wide test |
| 菌株编号 | 产量/(mg·mL-1) |
|---|---|
| SCSIO 17048 | 6.4533 |
| SCSIO 17023 | 6.4067 |
| SCSIO 17046 | 6.2300 |
| SCSIO 17003 | 6.1800 |
| SCSIO 17036 | 6.0767 |
| SCSIO 17037 | 6.0467 |
| SCSIO 17054 | 6.0200 |
| SCSIO 17111 | 5.9833 |
| SCSIO 17100 | 5.9733 |
| SCSIO 17096 | 5.9633 |
| SCSIO 17049 | 5.8067 |
| SCSIO 17060 | 5.6267 |
| SCSIO 17061 | 5.6233 |
| SCSIO 17019 | 5.5200 |
| SCSIO 17051 | 5.4067 |
| SCSIO 17076 | 5.4033 |
| SCSIO 17099 | 5.1933 |
| SCSIO 17097 SCSIO 17010 | 5.0767 5.0267 |
表3 菌株胞外多糖产量Tab. 3 Extracellular polysaccharide productions |
| 菌株编号 | 产量/(mg·mL-1) | 菌株来源 | 菌株属 |
|---|---|---|---|
| SCSIO 17111 | 0.0754 | 永暑礁结皮底层砂 | Lysobacter |
| SCSIO 17003 | 0.0539 | 永暑礁结皮层 | Bacillus |
| SCSIO 17096 | 0.0403 | 三亚结皮层 | Agromyces |
| SCSIO 17051 | 0.0307 | 三亚结皮层 | Bacillus |
| SCSIO 17100 | 0.0305 | 三亚结皮层 | Microbacterium |
| SCSIO 17049 | 0.0252 | 三亚结皮层 | Bacillus |
| SCSIO 17010 | 0.0213 | 永暑礁结皮层 | Bacillus |
| SCSIO 17061 | 0.0209 | 三亚结皮层 | Pseudoxanthomonas |
| SCSIO 17048 | 0.0186 | 三亚结皮层 | Nocardioides |
| SCSIO 17060 | 0.0185 | 三亚结皮层 | Devosia |
| SCSIO 17097 | 0.0182 | 三亚结皮层 | Brachybacterium |
| SCSIO 17023 | 0.0180 | 永暑礁结皮底层砂 | Bacillus |
| SCSIO 17046 | 0.0170 | 三亚结皮层 | Halobacillus |
| SCSIO 17076 | 0.0168 | 永暑礁结皮层 | Microbacterium |
| SCSIO 17099 | 0.0165 | 永暑礁结皮底层砂 | Bacillus |
| SCSIO 17036 | 0.0161 | 三亚结皮层 | Mesorhizobium |
| SCSIO 17037 | 0.0158 | 三亚结皮层 | Ornithinimicrobium |
| SCSIO 17054 | 0.0151 | 三亚结皮层 | Bacillus |
| SCSIO 17019 | 0.0137 | 永暑礁结皮底层砂 | Pseudomonas |
图5 SCSIO 17111菌株在粒径0.2~1.25mm(a、b)和<0.2mm (c、d)的珊瑚砂里的固砂实验效果Fig. 5 Sand fixation of strain SCSIO 17111 in fraction 0.2-1.25 mm (a, b) and < 0.2 mm (c, d) sand experimental |
表4 菌株固砂能力Tab. 4 Sand fixation capacity of strains |
| 菌株编号 | 土壤团聚体厚度/mm | 干筛结果/g (粒径>1.25mm) | 菌株来源 | 菌株属 |
|---|---|---|---|---|
| SCSIO 17111 | 6.00±0.13 | 13.56 | 永暑礁结皮底层砂 | Lysobacter |
| SCSIO 17023 | 3.89±0.14 | 0 | 永暑礁结皮底层砂 | Bacillus |
| SCSIO 17096 | 4.44±0.14 | 0 | 三亚结皮层 | Agromyces |
| SCSIO 17049 | 3.67±0.07 | 0 | 三亚结皮层 | Bacillus |
| SCSIO 17051 | 3.78±0.10 | 0 | 三亚结皮层 | Bacillus |
| SCSIO 17003 | 3.33±0.13 | 0 | 永暑礁结皮层 | Bacillus |
| SCSIO 17061 | 3.78±0.07 | 0 | 三亚结皮层 | Pseudoxanthomonas |
| SCSIO 17100 | 2.00±0.19 | 0 | 三亚结皮层 | Microbacterium |
| SCSIO 17010 | 2.11±0.20 | 0 | 永暑礁结皮层 | Bacillus |
| [1] |
艾雪, 2015. 沙漠结皮中耐盐碱细菌的分离及其固沙特性研究[D]. 兰州: 兰州交通大学.
|
| [2] |
艾雪, 王艺霖, 张威, 等, 2015. 柴达木沙漠结皮中耐盐碱细菌的分离及其固沙作用研究[J]. 干旱区资源与环境, 29(10): 145-151.
|
| [3] |
董群, 郑丽伊, 方积年, 1996. 改良的苯酚-硫酸法测定多糖和寡糖含量的研究[J]. 中国药学杂志, (9): 38-41.
|
| [4] |
毛龙江, 张永战, 魏灵, 等, 2006. 海南岛三亚湾海滩研究[J]. 第四纪研究, 26(3): 477-484.
|
| [5] |
南沙海域环境质量研究专题组, 1996. 南沙群岛及其邻近海域环境质量研究[M]. 北京: 海洋出版社: 1-111.
Nansha sea area environmental quality research theme group, 1996. The research on environment quality in the Nansha Islands and adjacent sea area[M]. Beijing: China Ocean Press: 1-111 (in Chinese).
|
| [6] |
钱琨, 王新志, 陈剑文, 等, 2017. 南海岛礁吹填钙质砂渗透特性试验研究[J]. 岩土力学, 38(6): 1557-1564.
|
| [7] |
任玉宾, 2016. 南海钙质砂渗透特性试验研究[D]. 大连: 大连理工大学.
|
| [8] |
王琳, 2020. 南海典型岛礁生物土壤结皮的微生物组及培育技术研究[D]. 广州: 中国科学院南海海洋研究所.
|
| [9] |
王新志, 2008. 南沙群岛珊瑚礁工程地质特性及大型工程建设可行性研究[D]. 武汉: 中国科学院研究生院 (武汉岩土力学研究所).
|
| [10] |
吴楠, 潘伯荣, 张元明, 2004. 土壤微生物在生物结皮形成中的作用及生态学意义[J]. 干旱区研究, (4): 444-450.
|
| [11] |
谢作明, 2006. 荒漠藻类对紫外辐射的响应及其结皮形成的研究[D]. 武汉: 中国科学院研究生院(水生生物研究所).
|
| [12] |
余劲聪, 2016. 海藻寡糖在农业领域的应用研究进展[J]. 南方农业学报, 47(6): 921-792.
|
| [13] |
张偲, 王琳, 李洁, 2018. 一株产胞外多糖可固沙的溶杆菌SCSIO 17111及其应用[P]. 广东: CN108795807A, 2018-11-13 (in Chinese).
|
| [14] |
张偲, 王琳, 李洁, 2021. 一种适用于南海珊瑚岛礁钙质砂土壤的藻菌结皮培育方法[P]. 广东: CN113287382A, 2021-08-24 (in Chinese).
|
| [15] |
张文平, 李昆太, 黄林, 等, 2017. 产胞外多糖菌株的筛选及其对土壤团聚体的影响[J]. 江西农业大学学报, 39(4): 772-729.
|
| [16] |
郑楠, 邵阳, 罗敏, 等, 2022. 土壤团聚体制备方法对其稳定性及固碳潜力评价的影响研究[J]. 中国环境科学, 42(6): 2821-2827.
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}