
南海北部涠洲岛边缘珊瑚礁的生物侵蚀实验研究
|
江绿苗(1998—), 女, 海南省三亚市人, 硕士研究生, 主要从事珊瑚礁沉积学研究。email: |
Copy editor: 林强
收稿日期: 2023-01-03
修回日期: 2023-03-26
网络出版日期: 2023-04-13
基金资助
国家自然科学基金(42076065)
中央级公益性科研院所基本科研业务专项(PM-zx703-202105-176)
中央级公益性科研院所基本科研业务专项(PM-zx703-202004-143)
Experimental study on bioerosion of marginal reefs in the Weizhou Island, northern South China Sea
Copy editor: LIN Qiang
Received date: 2023-01-03
Revised date: 2023-03-26
Online published: 2023-04-13
Supported by
National Natural Science Foundation of China(42076065)
Central Public-Interest Scientific Institution Basal Research Fund(PM-zx703-202105-176)
Central Public-Interest Scientific Institution Basal Research Fund(PM-zx703-202004-143)
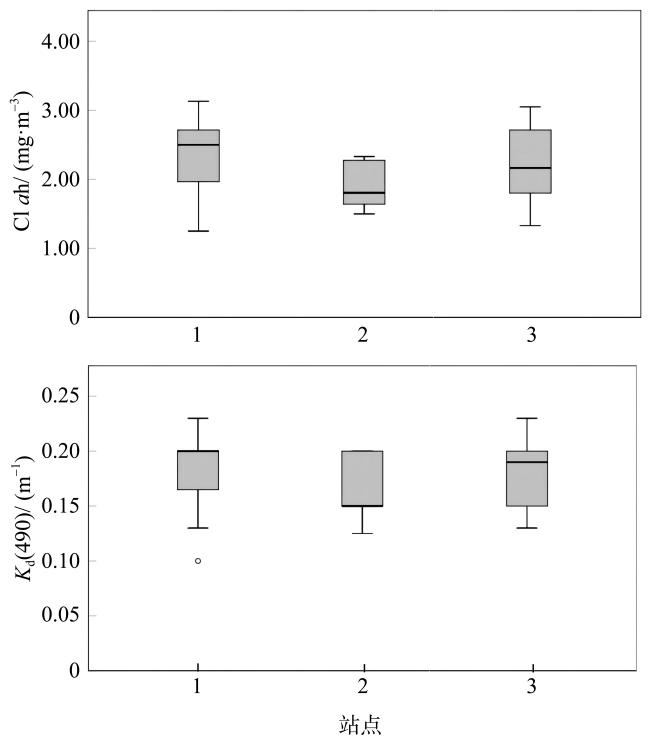
在全球变化背景下, 边缘珊瑚礁面临着来自自然与人类的双重压力。海水富营养化引起的生物侵蚀加剧, 虽然较为隐蔽, 但却会对珊瑚群落的发育以及礁格架的稳定性产生不良影响, 是边缘珊瑚礁潜在的危害之一。本研究在南海北部涠洲岛三个典型的边缘珊瑚礁开展了近一年的原位侵蚀实验, 定量分析了珊瑚礁的生物侵蚀强度与侵蚀速率, 并结合卫星遥感监测的水体环境参数, 探究了珊瑚礁侵蚀与海水环境的关系。研究发现, 内部大型侵蚀生物的侵蚀速率相比侵蚀强度更能够指示海水富营养盐与高浊度环境, 而侵蚀强度能够体现侵蚀生物的群落演替阶段。此外, 将涠洲岛生物侵蚀速率数据与全球其他典型珊瑚礁区域进行对比, 发现涠洲岛生物侵蚀速率处于全球中等水平(以内部大型侵蚀生物为主导), 但在其所属的“人为边缘礁”类型中为较低水平。结合气候变化、城市化发展、礁体健康状况等趋势以及本研究结果, 推测涠洲岛的生物侵蚀可能会继续加强, 达到“人为边缘礁”的高侵蚀水平, 并进一步加剧珊瑚礁的退化。
江绿苗 , 陈天然 , 赵宽 , 张婷 , 许莉佳 . 南海北部涠洲岛边缘珊瑚礁的生物侵蚀实验研究[J]. 热带海洋学报, 2024 , 43(3) : 155 -165 . DOI: 10.11978/2023002
In the context of global change, marginal coral reefs are threatened by both natural and anthropogenic disturbances. Biological erosion caused by seawater eutrophication is one of the potential hazards of marginal coral reefs, which can adversely impact the development of the coral community and the reef-framework stability. Here, in-situ bioerosion experiments were conducted for nearly a year on three typical marginal reefs in the Weizhou Island, northern South China Sea, to quantify coral reef bioerosion intensities and rates. Combined with the water environment parameters monitored by satellite remote sensing, we explored the relationship between coral reef erosion and water-quality parameters. We suggested that the bioerosion rates of internal bioeroders were more indicative of seawater eutrophication and high turbidity environment than the bioerosion intensities, which could reflect the community evolutionary stage of bioeroders. In addition, by comparing the bioerosion rate data of the Weizhou Island with other typical reef areas, we found that its bioerosion rate was at a moderate level (dominated by internal macroborers) in the world, but at a low level among the “anthropogenic marginal reefs”. Combined with the trends of climate change, urbanization development, reef health status, as well as the results of this study, we speculated that the biological erosion of the Weizhou Island may strengthen further to reach a high level for the “anthropogenic marginal reefs”, and may aggravate the degradation of coral reefs.
图2 原位侵蚀实验不同时期的珊瑚块体a. 原位侵蚀实验放置水下之前的珊瑚块(5×5 cm)与固定架; b—c. 回收清洗前的珊瑚块; d—i. 在各站点回收清洗后的珊瑚块的侵蚀痕迹(B: 双壳类; W: 蠕虫; S: 海绵)。图片左上角标注格式为“站点-样品编号” Fig. 2 The Coral blocks at different periods of the in-situ bioerosion experiment. (a) Coral blocks (5 × 5cm) with holders before placement underwater in the in-situ bioerosion experiment; (b-c) coral blocks before cleaning after recovery; (d-i) bioerosion traces of the coral blocks after recycling at each site (B, bivalves; W, worms; S, sponges). The upper left corner of these figures is labeled with the format "Site - Sample number" |
表1 内部大型侵蚀生物的侵蚀强度与侵蚀速率Tab. 1 Bioerosion intensity and rate of each internal macro-bioeroder |
| 站点 | 侵蚀速率/(kg·m-2·yr-1) | 侵蚀强度/% | 总侵蚀强度/% | ||
|---|---|---|---|---|---|
| 双壳类 | 海 绵 | 蠕 虫 | |||
| 1 | 0.60±0.16 | 0.09±0.06 | 0.15±0.15 | 0.56±0.30 | 0.80±0.44 |
| 2 | 0.45±0.15 | 0.05±0.05 | 0.03±0.02 | 0.24±0.05 | 0.32±0.08 |
| 3 | 0.38±0.11 | 0.03±0.03 | 0.08±0.05 | 0.29±0.05 | 0.40±0.09 |
| 涠洲岛(平均) | 0.48±0.07 | 0.06±0.05 | 0.09±0.15 | 0.38±0.10 | 0.52±0.25 |
注: 数据格式为平均值±标准差 |
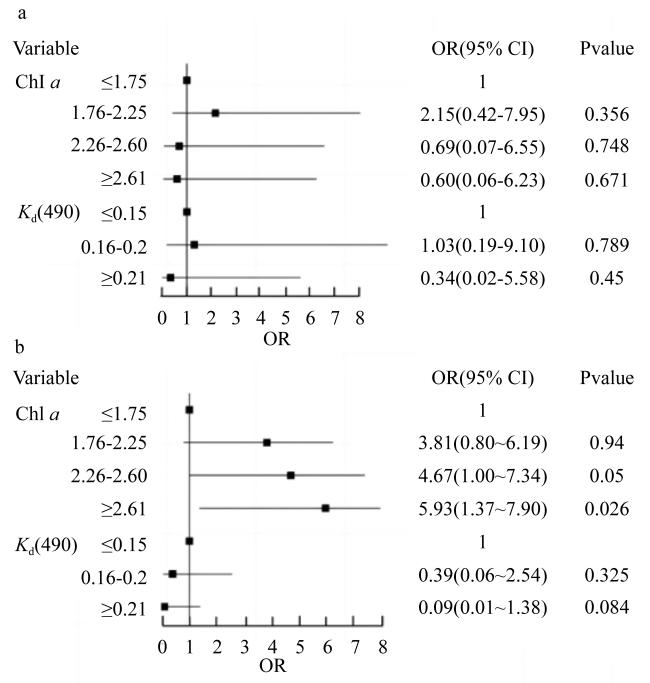
图4 生物侵蚀强度(a)和生物侵蚀速率(b)与水体环境参数的广义线性模型OR(odds ratio)表示优势比, 反映的是某种因素(本文是生物侵蚀强度和生物侵蚀速率)与因变量[本文是Chl a和Kd(490), 单位分别为mg·m-3和m-1]的关联强度。OR<1, 自变量起抑制作用; OR=1时, 自变量与因变量没有相关性; OR>1时, 自变量起促进作用, 并且OR越大, 促进作用越显著 Fig. 4 Plots of the GML model used for assessing the impacts of water quality parameters on the bioerosion intensity (a) and the bioerosion rate (b) of macroborers |
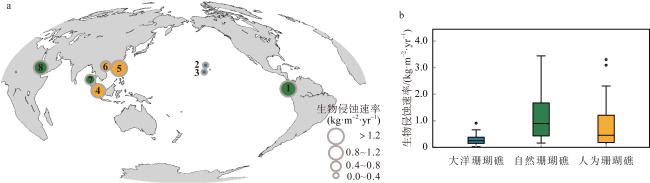
表2 全球珊瑚礁的生物侵蚀原位实验的研究结果Tab. 2 The results of in-situ experiments of bioerosion in the global coral reefs |
| 海域 | 地点 | 序号 | 暴露时间/月 | 生物侵蚀速率/(kg·m-2·yr-1) | 主要侵蚀生物 | 海水Chl a /(mg·m-3) | 海水Kd(490)/(m-1) | 区域特征 | 参考文献 |
|---|---|---|---|---|---|---|---|---|---|
| 东热带太平洋 | 巴拿马 | ① | 24 | 1.36 | 双壳类、蠕虫 | 1.99 | 0.16 | 自然边缘礁 | Enochs et al, 2021 |
| 中太平洋 | 茂纳卢阿湾 (夏威夷欧胡岛) | ② | 12 | 0.04 | (未知) | 0.09 | 0.00 | 大洋珊瑚礁 | Lubarsky et al, 2018 |
| 卡内奥赫湾 (夏威夷欧胡岛) | ③ | 24 | 0.34 | (未知) | 0.20 | 0.00 | 大洋珊瑚礁 | Silbiger et al, 2016 | |
| 西热带太平洋 | 新加坡南部岛屿 | ④ | 12 | 1.08 | 海绵、蠕虫 | 2.85 | 0.25 | 人为边缘礁 | Januchowski-Hartley et al, 2020 |
| 中国香港 | ⑤ | 24 | 1.38 | 海胆、 蠕虫、双壳类 | 3.16 | 0.19 | 人为边缘礁 | Yeung et al, 2021 | |
| 中国涠洲岛 | ⑥ | 10 | 0.46 | 蠕虫 | 2.19 | 0.19 | 人为边缘礁 | ||
| 印度洋 | 泰国斯米兰群岛 (安达曼海) | ⑦ | 18 | 0.70 | 鹦嘴鱼 | 0.25 | 0.04 | 自然边缘礁 | Schmidt et al, 2013 |
| 红海 | 沙特阿拉伯 中部海湾 | ⑧ | 30 | 0.96 | 内部大型生物 | 0.76 | 0.05 | 自然边缘礁 | Roik et al, 2018 |
图5 全球珊瑚礁的生物侵蚀原位实验地点(a)及研究结果(b)a. ①巴拿马海湾; ②茂纳卢阿湾(夏威夷欧胡岛); ③卡内奥赫湾(夏威夷欧胡岛); ④新加坡南部岛屿; ⑤中国香港; ⑥中国涠洲岛; ⑦泰国斯米兰群岛(安达曼海); ⑧沙特阿拉伯中部海湾 Fig. 5 Results of in situ experiments of bioerosion from global coral reefs. (a) ① Panama Bay; ② Maunalua Bay, O’ahu; ③ Kāne'ohe Bay, Hawai'i; ④ southern islands of Singapore; ⑤ HongKong, China; ⑥Weizhou Island, China; ⑦ Similan Islands (Thailand) in the Andaman Sea; ⑧ Saudi Arabian central Red Sea |
| [1] |
陈天然, 郑兆勇, 莫少华, 等, 2013. 涠洲岛滨珊瑚中的生物侵蚀及其环境指示意义[J]. 科学通报, 58(17): 1574-1582.
|
| [2] |
何精科, 黄振鹏, 2019. 广西涠洲岛珊瑚分布状况研究[J]. 海洋开发与管理, 36(1): 57-62.
|
| [3] |
黄晖, 马斌儒, 练健生, 等, 2009. 广西涠洲岛海域珊瑚礁现状及其保护策略研究[J]. 热带地理, 29(4): 307-312, 318.
|
| [4] |
黄晓煦, 徐轶肖, 张腾, 等, 2021. 涠洲岛海域营养盐变化特征与评价[J]. 广西科学, 28(2): 130-135.
|
| [5] |
金昱昕, 陈天然, 孟庆山, 等, 2017. 单轴抗压强度揭示南海珊瑚骨骼结构的差异[J]. 热带海洋学报, 36(2): 33-39.
|
| [6] |
梁文, 黎广钊, 张春华, 等, 2010. 20年来涠洲岛珊瑚礁物种多样性演变特征研究[J]. 海洋科学, 34(12): 78-87.
|
| [7] |
梁文, 张春华, 叶祖超, 等, 2011. 广西涠洲岛造礁珊瑚种群结构的空间分布[J]. 生态学报, 31(1): 39-46.
|
| [8] |
汤超莲, 周雄, 郑兆勇, 等, 2013. 未来海平面上升对涠洲岛珊瑚礁的可能影响[J]. 热带地理, 33(2): 119-123, 140.
|
| [9] |
张文静, 郑兆勇, 张婷, 等, 2020. 1960-2017年北部湾珊瑚礁区海洋热浪增强原因分析[J]. 海洋学报, 42(5): 41-48.
|
| [10] |
赵宽, 张婷, 陈天然, 2019. 南海北部滨珊瑚骨骼微生物侵蚀[J]. 热带海洋学报, 38(6): 74-79.
|
| [11] |
周浩郎, 黎广钊, 2014. 涠洲岛珊瑚礁健康评估[J]. 广西科学院学报, 30(4): 238-247.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}