
牙鲆(Paralichthys olivaceus)抗缪勒氏管激素Ⅱ型受体基因(amhr2)表达特征分析及功能初探
|
李泽 (1998—), 男, 山东青岛人, 硕士研究生, 主要从事鲆鲽鱼类发育生物学研究。email: |
Copy editor: 孙翠慈
收稿日期: 2023-03-24
修回日期: 2023-04-21
网络出版日期: 2023-05-25
基金资助
国家重点研发计划(2022YFD2400402)
国家重点研发计划(2018YFD0900202)
山东省自然科学基金(ZR2022MC026)
青岛海洋科学与技术试点国家实验室海洋生物学与生物技术功能实验室青年科学基金项目(YQ2018NO01)
Expression characteristics and function of Anti-Müllerian hormone receptor Ⅱ gene (amhr2) in Paralichthys olivaceus
Copy editor: SUN Cuici
Received date: 2023-03-24
Revised date: 2023-04-21
Online published: 2023-05-25
Supported by
National Key R&D Program of China(2022YFD2400402)
National Key R&D Program of China(2018YFD0900202)
Shandong Natural Science Foundation(ZR2022MC026)
Youth Research Fund of Marine Biology and Biotechnology Laboratory, Pilot National Laboratory for Marine Science and Technology (Qingdao)(YQ2018NO01)
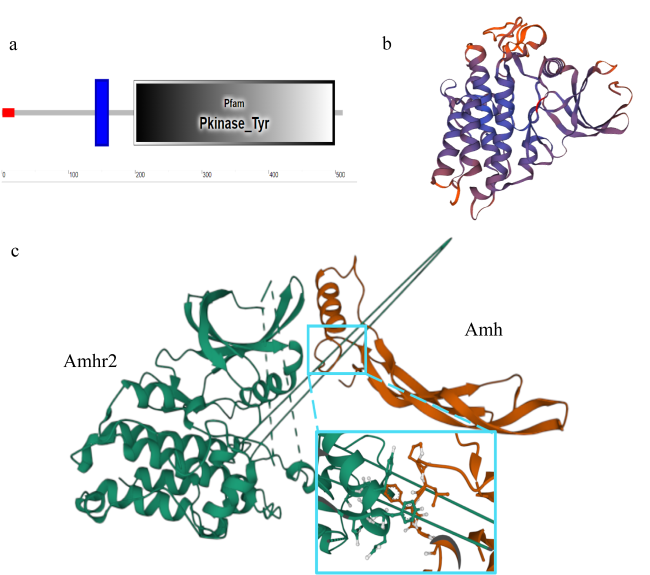
抗缪勒氏管激素Ⅱ型受体(anti-Müllerian hormone receptor Ⅱ, Amhr2)是抗缪勒氏管激素(anti-Müllerian hormone, Amh)的特异受体。amhr2基因在鱼类性腺分化和发育中发挥重要作用, 更决定了有的鱼种的性别, 然而, 相关功能研究较为有限。本研究旨在明晰amhr2在我国重要海水养殖鱼类—牙鲆(Paralichthys olivaceus)性腺分化和发育过程中的表达特征, 并初步探究其功能。首先, 克隆了牙鲆amhr2-CDS序列, 共1536bp, 编码511个氨基酸。系统发育分析显示牙鲆Amhr2近C端较为保守, 与其他硬骨鱼类聚为一枝, 并与上游sp1以及下游的prr13和PCBP2在基因组中共定位。蛋白结构分析显示, 牙鲆Amhr2包含信号肽、跨膜结构域和保守的酪氨酸蛋白激酶结构域。进而, 利用实时荧光定量PCR (qPCR)分析表明, 牙鲆amhr2主要表达于性腺, 且在精巢中的表达极显著高于卵巢(P< 0.01), 其在I-V期精巢中持续高表达, 而在卵巢中仅在I期高表达, 后显著下降(P< 0.05)。性腺分化期基因的表达, 是对雌核发育组(对照组, 20 ± 0.5℃, 100%雌性)与雌核发育高温诱导组(HT组, 28 ± 0.5℃, 100%雄性)鱼苗进行检测的, amhr2在对照组全长(total length, TL)2cm的实验鱼性腺中表达最高, 后逐渐下降; 而HT组实验鱼性腺中的表达呈现先上升后下降的趋势, 并在精巢开始分化的6cm TL时表达最高。利用Hela细胞进行亚细胞定位分析发现, Amhr2与其配体Amh均定位于细胞质。同时, 通过原核表达重组牙鲆Amh, 并用其孵育离体性腺组织, qPCR分析显示精巢中amhr2的表达没有显著变化, 但卵巢中表达显著上升(P< 0.05)。进一步通过双荧光素酶报告分析表明, Amh和Amhr2共转染能够显著抑制雌激素合成的关键芳香化酶基因cyp19a的表达(P< 0.01)。综上所述, 牙鲆amhr2主要表达于精巢, 但在不同性腺发育时期表达不同, 且其可能与Amh共同影响cyp19a的转录, 对雄性表型形成的启动和性腺发育起作用。
关键词: 牙鲆(Paralichthys olivaceus); amhr2; 性腺分化和发育; 基因表达; 功能
李泽 , 王丽娟 , 邹聪聪 , 舒畅 , 吴志昊 , 邹玉霞 , 尤锋 . 牙鲆(Paralichthys olivaceus)抗缪勒氏管激素Ⅱ型受体基因(amhr2)表达特征分析及功能初探[J]. 热带海洋学报, 2024 , 43(1) : 94 -106 . DOI: 10.11978/2023040
Anti-Mullerian hormone receptor Ⅱ (Amhr2) is the specific receptor of Anti-Mullerian hormone (Amh) in the TGF-β pathway. It is critical for gonadal differentiation and development in fish and determination of the sex in some fish species. However, the expression and function of amhr2 in fish has been poorly studied. To clarify the expression characteristics and function of amhr2, a 1536 bp CDS encoding 511 amino acids, was obtained from Paralichthys olivaceus, an important commercially cultured marine fish in China. The flounder Amhr2 protein was clustered with those from other teleosts, with the highly conservative motifs occurring in the C-terminal region. Gene collinearity analysis revealed that the amhr2 collaborates with specificity protein 1 (sp1) upstream, and proline-rich protein 13 (prr13) and poly (rC) binding protein 2 (PCBP2) downstream of the teleost genome. The predicted flounder Amhr2 protein was composed of a signal peptide, two transmembrane domains, and a conservative tyrosine protein kinase domain. Real-time quantitative PCR (qPCR) results showed that amhr2 was mainly expressed in the adult gonads, with significantly higher expression in the testes than in the ovaries (P<0.01). At stages I-V of the testis, the expression of amhr2 remained at a high level, whereas the expression level in the ovaries was significantly higher at stage I than at stages Ⅱ-V (P< 0.05). During the flounder gonadal differentiation period, gynogenesis (control, 20 ± 0.5℃, 100% female) and gynogenesis at high temperature (HT, 28 ± 0.5℃, 100% male) were used to detect the expression of amhr2. The results showed that the expression of amhr2 in the control group was highest at 2 cm total length (TL) and then decreased continuously, while the expression in the HT group first increased and then decreased. The inflection point of amhr2 expression in the HT group was at 6 cm TL, with the highest level. To investigate the effect of Amh on amhr2, we examined the co-localization of Amhr2 and Amh with Hela cells. Examination of subcellular localization showed that both Amhr2 and Amh were mainly located in the cytoplasm. Subsequently, the recombinant protein Amh of the flounder was obtained by prokaryotic expression. After in vitro incubation with the recombinant Amh, the expression of amhr2 in the testes showed no significant difference, whereas the transcription level in the ovaries increased significantly at 12 and 48 h (P<0.05). In addition, a dual luciferase reporter assay in HEK293T cells showed that co-transfection of Amh and Amhr2 could inhibit the promoter activity of the cytochrome P450 aromatase gene (cyp19a) (P<0.01), the key gene for estrogen synthesis. These findings suggest that amhr2 is mainly expressed in the testis, but its expression varies at different stages of the gonadal development. And amhr2 may affect the transcription of cyp19a together with Amh, which plays a role in the onset of the male phenotype formation and gonadal development in the flounder.
表1 本研究所用引物Tab. 1 Primers used in this study |
| 引物 | 序列 | NCBI序列号及参考文献 |
|---|---|---|
| amhr2-CDS-F | ATTCCCAGGACCCTCCCTGTT | XM_020096590.1 |
| amhr2-CDS-R | TCTGCAAAGAGAACAGTGCAAC | |
| amh-CDS-F | ATGCCGGTGGTGAACGTCTT | XM_020080898.1 |
| amh-CDS-R | GCGGCATCCACACTCCTTTG | |
| amhr2-qPCR-F | CGTCGTGTTCCTGCCAAAC | XM_020096590.1 |
| amhr2-qPCR-R | GGCAGCTCATACACCTCCTT | |
| actin- qPCR-F | GGAATCCACGAGACCACCTACA | EU090804 Hu et al, 2014 |
| actin- qPCR-R | CTGCTTGCTGATCCACATCTGC | |
| ef- qPCR-F | AGCCAGAAGCGTTTTGAGGAG | XM_020104638.1 Zhong et al, 2008 |
| ef- qPCR-R | AGATGGGGACGAAAGCAACAC | |
| amh-pc3.1-EcoRV-F | CACACTGGACTAGTGGATCCGCCACCATGCCGGTGGTGAACGTC | XM_020080898.1 |
| amh-pc3.1-EcoRV-R | AGCTTGGTACCGAGCTCGGATCCGCGGCATCCACACTCCT | |
| amhr2-pc3.1-BamHI-F | CACACTGGACTAGTGGATCCGCCACCATGATGCTGCAGTGGTGGC | XM_020096590.1 |
| amhr2-pc3.1-BamHI-R | AGCTTGGTACCGAGCTCGGATCCTGAGCTGCTTTCAGAGACATAAGACTG | |
| amhr2-pCS2-BamHI-F | ACGACGACGATAAGGGATCCGCCACCATGATGCTGCAGTGGTGGC | XM_020096590.1 |
| amhr2-pCS2-BamHI-R | AATTCGAATCGATGGGATCCTGAGCTGCTTTCAGAGACATAAGACTG | |
| amh-pET-EcoRV-F | AAGGCCATGGCTGATATGCCGGTGGTGAACGTCTT | XM_020080898.1 |
| amh-pET-EcoRV-R | GAATTCGGATCCGATGCGGCATCCACACTCCTTTG |
表2 细胞转染组别及组分Tab. 2 Cell transfection groups and their components |
| 转染组分(ng)\组别 | cyp19a转染对照组 | cyp19a+amh共转染组 | cyp19a+amhr2共转染组 | cyp19a+amh+amhr2共转染组 |
|---|---|---|---|---|
| cyp19a-pGL3 | 400 | 400 | 400 | 400 |
| amh-pc3.1 | 0 | 400 | 0 | 200 |
| amhr2-pc3.1 | 0 | 0 | 400 | 200 |
| TK-pRL | 40 | 40 | 40 | 40 |
| pcDNA 3.1 | 560 | 160 | 160 | 160 |
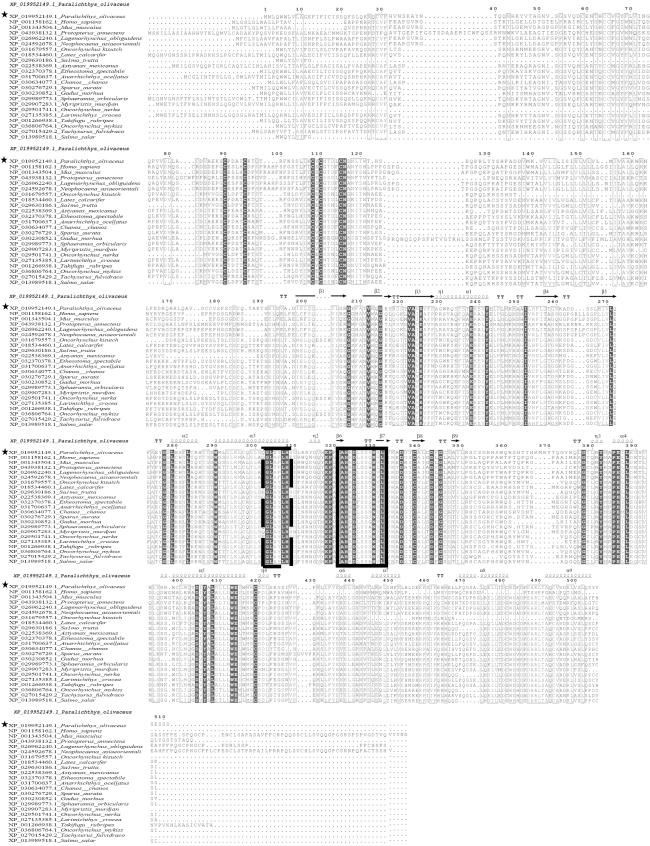
图1 牙鲆与其他物种Amhr2氨基酸序列多重比对及二级结构分析星号, 牙鲆; α, α-螺旋结构; β, β-折叠结构; 黑色区域, 序列完全相同区域; 虚线框, GL××LH保守基序; 实线框, AH×D××××N×L保守基序 Fig. 1 Amino acid sequence alignment and secondary structure prediction of Amhr2 among Paralichthys olivaceus and other species. P. olivaceus is marked with star. α, α- helices structure; β, β- sheets structure; black area, the sequence completely conserved region; dotted line box, the GL ×× LH conservative motif; full line box, the AH × D ×××× N × L conservative motif |
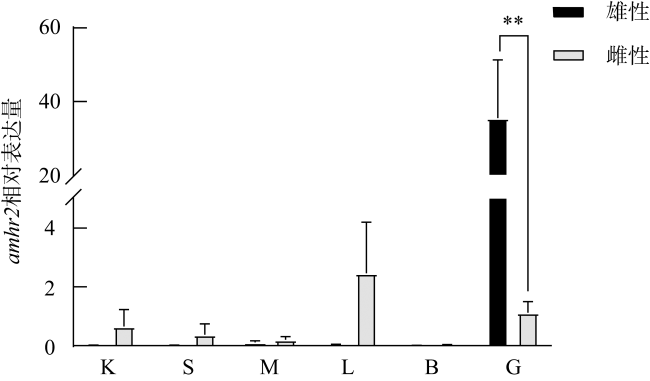
图4 牙鲆amhr2雌雄个体组织差异表达K, 肾; S, 脾; M, 肌肉; L, 肝; B, 脑; G, 性腺; n=3; **表示雌雄性腺具有极显著差异, P< 0.01 Fig. 4 The expression of amhr2 in different tissues of male and female P. olivaceus. K, kidney; S, spleen; M, muscle; L, liver; B, brain; G, gonad. n=3. ** indicates the difference between the testes and ovaries, P< 0.01 |
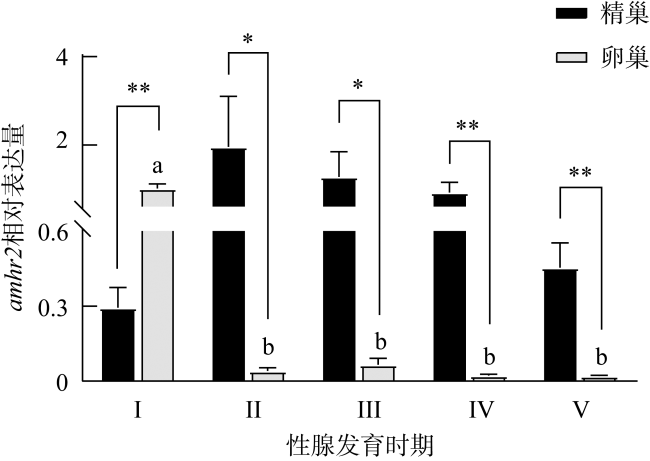
图5 牙鲆amhr2在性腺发育各时期的表达n=3。不同字母表示不同性腺发育时期卵巢中amhr2的表达具有显著差异, P< 0.05。*表示同一性腺发育时期精巢、卵巢中具有显著差异, P< 0.05; **, P< 0.01 Fig. 5 The expression of amhr2 in P. olivaceus during the gonadal development. n=3. Different letters indicate the difference in the ovaries of the fish at different gonadal development stages. * indicates the difference between the testes and ovaries of the fish at the same gonadal development stages, P< 0.05; **, P< 0.01 |
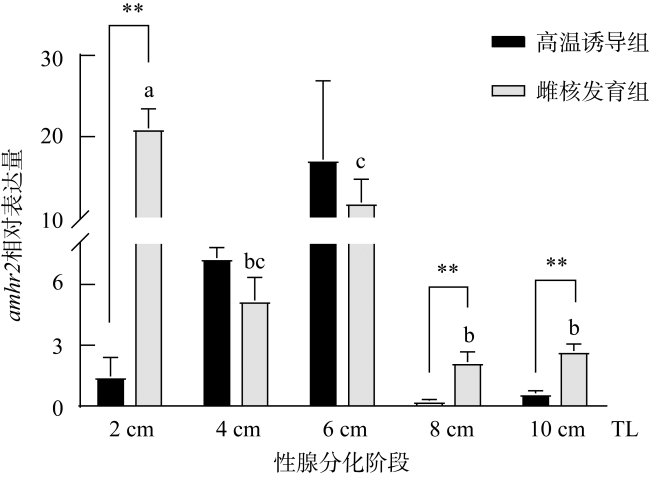
图6 牙鲆amhr2在性腺分化期的表达n=3。不同字母表示不同全长雌核发育对照组牙鲆性腺中amhr2表达具有显著差异, P< 0.05。**表示在相同全长时精巢、卵巢中具有极显著差异, P< 0.01 Fig. 6 The expression of amhr2 in P. olivaceus during gonadal differentiation. n=3. Different letters indicate the differences at different TLs of the gynogenesis control group during the gonadal differentiation, P< 0.05. ** indicates the difference between the testes and ovaries of the fish at the same TLs, P< 0.01 |
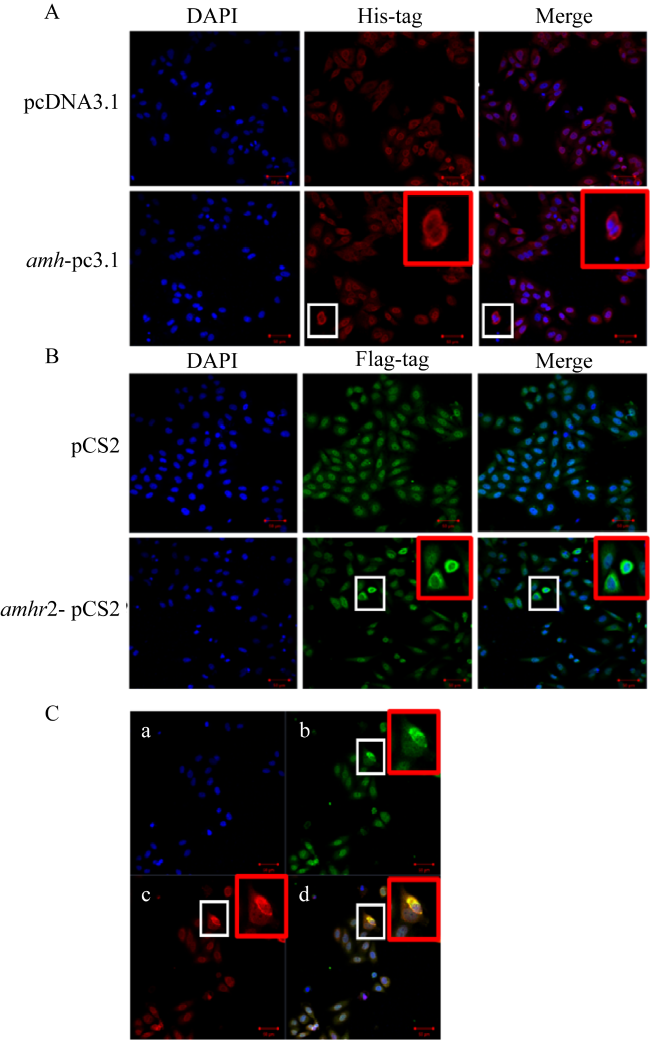
图7 牙鲆Amh和Amhr2的亚细胞定位A, Amh; pcDNA3.1, 空载体质粒; amh-pc3.1, amh重组质粒; B, Amhr2; pCS2, 空载体质粒; amhr2-pCS2, amhr2重组质粒; C, Amh和Amhr2的共定位 (a. DAPI; b. anti-Flag; c. anti-His; d. anti-Flag+ anti-His); 红色框, 白色框放大后图像, bar=50 μm Fig. 7 Subcellular localization of Amh and Amhr2 of P. olivaceus. (a) Amh, pcDNA3.1, blank vector plasmid; amh-pc3.1, recombinant plasmid of amh; (B) Amhr2, pCS2, blank vector plasmid; amhr2-pCS2, recombinant plasmid of amhr2; (c) the co-localization of Amh and Amhr2 (a. DAPI; b. anti-Flag; c. anti-His; d. anti-Flag+ anti-His). Red box, the enlarged image of the white box, bar=50 μm |
图8 牙鲆Amh蛋白的原核表达及纯化a. Amh原核表达SDS-PAGE结果(M, marker; 1, 空白培养基; 2, 实验培养基; 3, 空载菌体沉淀; 4, Amh菌体沉淀; 5, 空白菌体上清; 6, Amh菌体上清); b. 重组Amh纯化SDS-PAGE结果(M, marker; 1-7, 洗脱目的蛋白); c. Amh原核表达WB检测(M, marker; 1. 空白培养基; 2. 菌体沉淀; 3. 菌体破碎后上清; 4. 菌体破碎后沉淀;框, 重组Amh)。 Fig.8 Expression and purification of the recombinant P. olivaceus Amh. (a) SDS-PAGE results of Amh prokaryotic expression (M, marker; 1, blank medium; 2, experimental medium; 3, control thallus precipitation; 4, Amh thallus precipitation; 5, blank cell supernatant; 6, Amh cell supernatant); (b) SDS-PAGE results of recombinant Amh purification (M, marker; 1-7, protein elution in each step); (c) WB results of Amh prokaryotic expression (M, marker; 1, blank medium; 2, Amh thallus precipitation; 3, supernatant after the bacterial fragmentation; 4, precipitation after the bacterial fragmentation. Box, the recombinant Amh) |
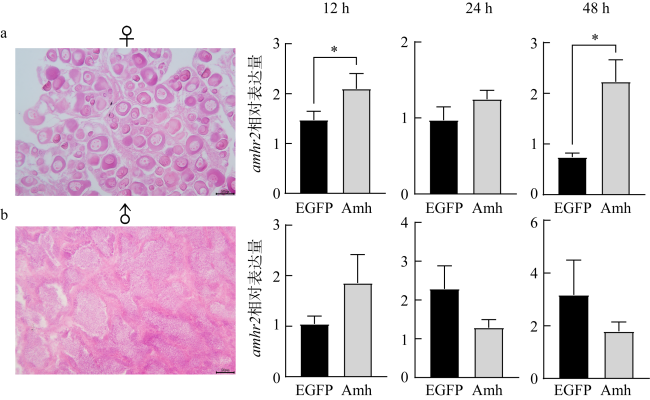
图9 重组Amh孵育后雌雄性腺中amhr2的表达变化a. 卵巢组织切片及孵育12、24、48h后amhr2表达变化; b. 精巢组织切片及孵育12、24、48h后amhr2表达变化。n=4; bar=50μm; *表示不同孵育组间具有差异, P< 0.05。 Fig. 9 The changes of amhr2 expression in the ovary and testis incubated with the recombinant Amh. (a) Histological section of the ovary and the expression of amhr2 after 12, 24, and 48 h incubation with recombinant Amh in the ovaries; (b) histological observation of the testis and the expression of amhr2 after 12, 24, 48 h incubation with the recombinant Amh in the testes. n=4. bar=50μm. * indicates the difference between different groups, P< 0.05 |
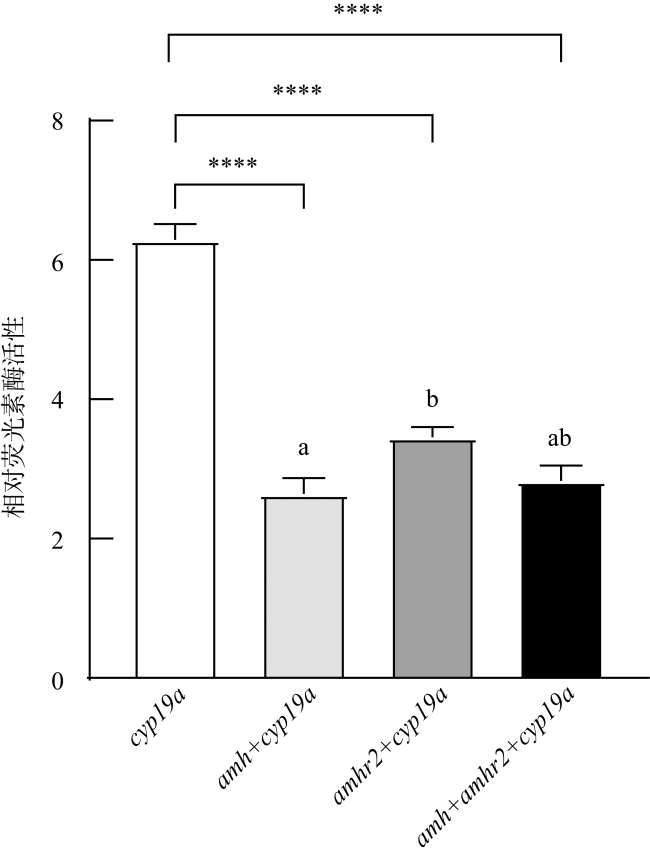
图10 牙鲆Amh与Amhr2对cyp19a的调控不同字母代表各实验组间存在显著差异, P< 0.05。****表示对照组与各实验组间存在极显著差异, P< 0.0001 Fig. 10 The regulation of Amh and Amhr2 on cyp19a in P. olivaceus. Different letters represent significant differences among the experimental groups, P< 0.05. **** indicates significant difference between the control group and experimental groups, P< 0.0001 |
| [1] |
范兆飞, 2017. 牙鲆cyp19a及其转录因子表观修饰和调控研究[D]. 青岛: 中国科学院大学: 中国科学院海洋研究所: 1-137.
|
| [2] |
高莹莹, 胡鹏, 刘新富, 等, 2019. 暗纹东方鲀抗苗勒氏管激素Ⅱ型受体基因的克隆、生物信息学及表达分析[J]. 海洋渔业, 41(5): 555-566.
|
| [3] |
韩玉龙, 2019. amh基因在斜带石斑鱼性别分化中的作用机制研究[D]. 广州: 中山大学: 1-130.
|
| [4] |
孙鹏, 尤锋, 张立敬, 等, 2009. 牙鲆性腺分化的组织学研究[J]. 海洋科学, 33(3): 53-58.
|
| [5] |
王妹, 邓思平, 陈华谱, 等, 2018. 金钱鱼Amhr2基因的克隆及表达分析[J]. 广东海洋大学学报, 38(3): 17-24.
|
| [6] |
赵九娥, 2015. Amhy/Amh及其受体AmhrⅡ对尼罗罗非鱼雄性性别的决定作用[D]. 重庆: 西南大学: 1-80.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}