
两种造礁石珊瑚固碳能力初步研究*
*感谢海南省财政项目蓝碳研究及应用示范(46000023T000000939334)和海南省重点研发项目(ZDYF2020177)的支持。
|
赵贺(1996—), 男, 山东省济宁市人, 博士研究生, 从事修复生态学研究。email: |
Copy editor: 林强
收稿日期: 2023-04-26
修回日期: 2023-06-13
网络出版日期: 2023-06-21
基金资助
海南省财政项目蓝碳研究及应用示范(46000023T000000939334)
海南省重点研发项目(ZDYF2020177)
Preliminary study on carbon sequestration capacity of two hermatypic corals*
Copy editor: LIN Qiang
Received date: 2023-04-26
Revised date: 2023-06-13
Online published: 2023-06-21
Supported by
Hainan Provincial Financial Project Blue Carbon Research and Application Demonstration(46000023T000000939334)
Hainan Province Key R&D Project(ZDYF2020177)
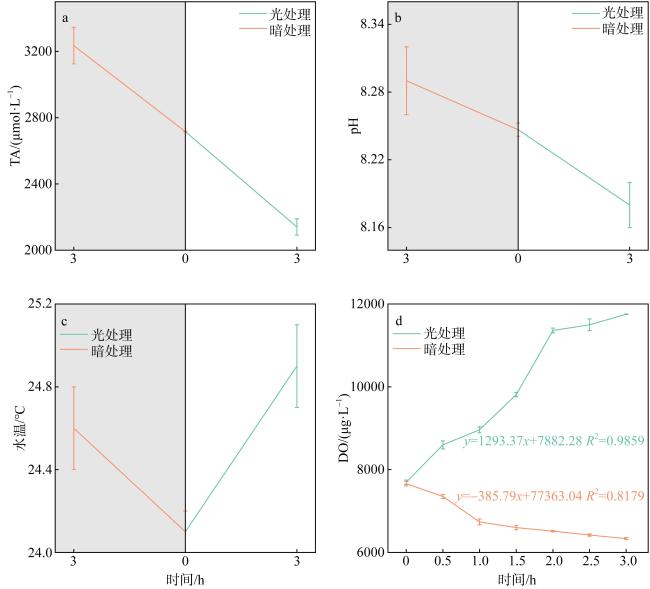
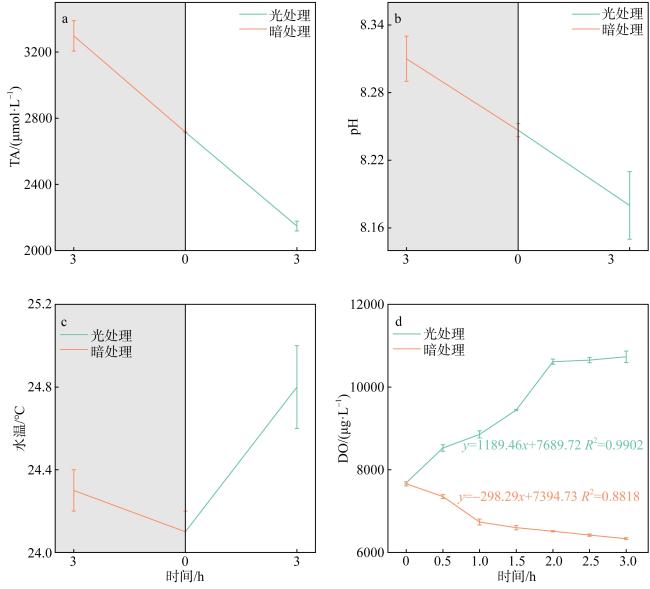
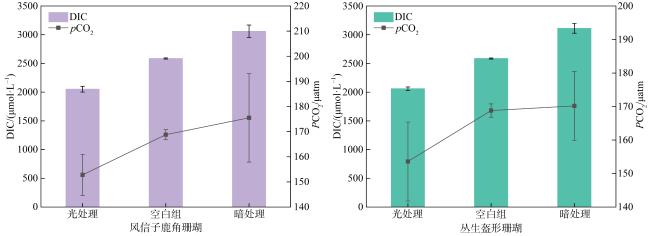
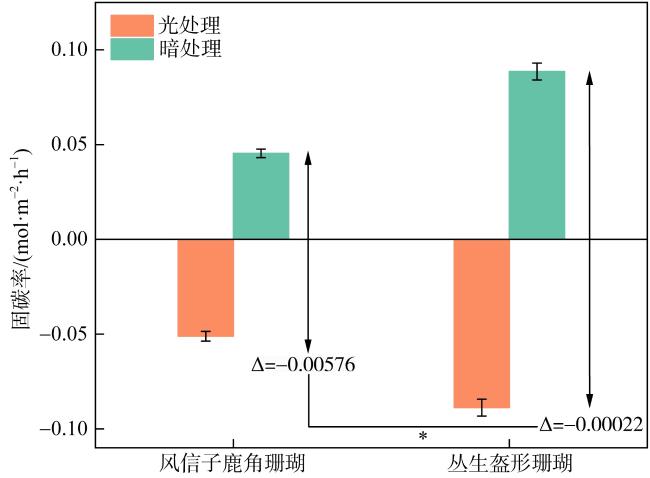
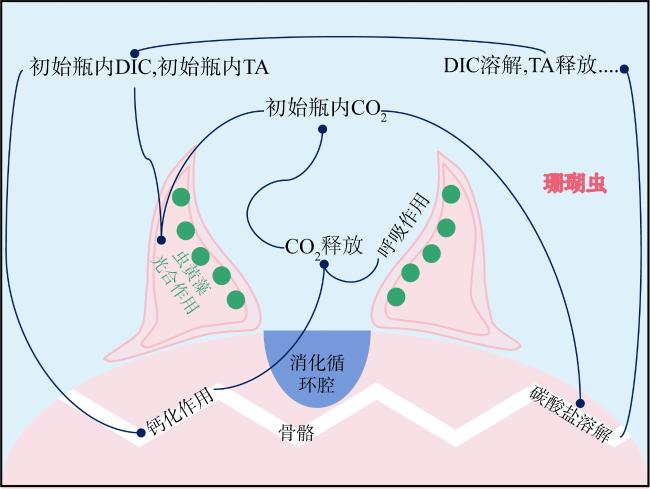
珊瑚礁是海洋中生产力水平最高的生态系统之一, 其碳循环受珊瑚光合作用、呼吸作用和钙化作用三大代谢过程的共同调节, 过程十分复杂。为探明珊瑚固碳能力, 明确其在光照和黑暗下固碳能力的变化过程, 本研究以取自三亚蜈支洲岛健康风信子鹿角珊瑚(Acropora hyacinthus)和丛生盔形珊瑚(Galaxea fascicularis)为研究对象, 基于碳酸盐体系分析方法, 通过设置光照、黑暗两种处理方式, 测定试验前后水体理化因子变化, 并根据CO2体系计算软件得出水体溶解无机碳(dissolved inorganic carbon, DIC)和二氧化碳分压(partial pressure of carbon dioxide, pCO2)含量。结果显示: 在光照条件下, 两种珊瑚所处水体总碱度(total alkalinity, TA)和酸碱度(pH)下降, 水温和溶解氧(dissolved oxygen, DO)上升, DIC和CO2被持续消耗, 产生碳汇效应; 黑暗条件下, 水体TA、pH和温度上升, DO下降, DIC和CO2持续释放, 产生碳源效应; 比较光暗处理结果后, 计算得出风信子鹿角珊瑚固碳率为0.00576mol·m-2·h-1; 丛生盔形珊瑚固碳率为0.00022mol·m-2·h-1, 风信子鹿角珊瑚固碳效率显著高于丛生盔形珊瑚(P<0.05); 两种珊瑚光合作用吸收CO2的量大于自身所产生的, 属于CO2的汇。综上所述, 两种珊瑚最终降低了水体TA、DIC和pCO2, 能够促进大气CO2向海水转移, 形成碱度汇和碳汇效应。
赵贺 , 张峻菱 , 王浩 , 柯景召 , 朱铭 , 王爱民 , 李秀保 . 两种造礁石珊瑚固碳能力初步研究*[J]. 热带海洋学报, 2024 , 43(3) : 146 -154 . DOI: 10.11978/2023053
Coral reefs stand as remarkably productive ecosystems within the vast ocean, wherein their carbon cycle is meticulously regulated through a complex interplay of three pivotal metabolic processes: photosynthesis, respiration, and calcification. These intricate mechanisms contribute to the intricate and sophisticated functioning of these ecosystems, rendering them worthy subjects of extensive scholarly inquiry and analysis. To explore the carbon sequestration potential of corals and shed light on the underlying mechanisms in both illuminated and dark conditions, this research was conducted using Acropora hyacinthus and Galaxea fascicularis specimens collected from the Wuzhizhou Island, Sanya. Employing the carbonate system, an analysis was carried out to assess the carbon sequestration capabilities of these two coral species. This involved measuring the changes in physicochemical factors in the surrounding water before and after the experiment, as well as determining the content of dissolved inorganic carbon (DIC) and partial pressure of carbon dioxide (pCO2) using CO2 system calculation software. By employing these methodologies, a comprehensive understanding of the carbon sequestration processes within these coral species was achieved. The findings of this study demonstrated distinct responses of the water parameters for A. hyacinthus and G. fascicularis under light and dark conditions. In the presence of light, the water bodies surrounding both corals exhibited a decrease in total alkalinity (TA) and pondus hydrogenii (pH), an increase in water temperature (WT) and dissolved oxygen (DO), as well as continuous consumption of DIC and CO2, resulting in a pronounced carbon sink effect. Conversely, under dark conditions, TA, pH, and WT increased, DO decreased, and DIC and CO2 were consistently released, indicating a carbon source effect. By comparing the outcomes of light and dark treatments, the carbon sequestration rate of A. hyacinthus was calculated to be 0.00576 mol·m-2·h-1, while that of G. fascicularis was determined to be 0.00022 mol·m-2·h-1. Notably, the carbon sequestration efficiency of A. hyacinthus surpassed that of G. fascicularis significantly (P<0.05). Both corals exhibited a net absorption of CO2 through photosynthesis, surpassing the amount they produced, thereby acting as CO2 sinks. In summary, these two coral species effectively reduced the total alkalinity, DIC, and pCO2 within the water column, facilitating the transfer of atmospheric CO2 to seawater and engendering alkalinity sink and carbon sink effects.
| [1] |
黄建中, 魏宇衡, 顾志峰, 等, 2020. 海南西岛珊瑚群落变化及其影响因素[J]. 热带海洋学报, 39(6): 103-113.
|
| [2] |
刘小菊, 施祺, 陶士臣, 等, 2022. 近165年来中沙环礁中北暗沙滨珊瑚生长率及其对海温变化的响应[J]. 热带海洋学报, 41(5): 64-73.
|
| [3] |
龙丽娟, 杨芳芳, 韦章良, 2019. 珊瑚礁生态系统修复研究进展[J]. 热带海洋学报, 38(6): 1-8.
|
| [4] |
石拓, 郑新庆, 张涵, 等, 2021. 珊瑚礁: 减缓气候变化的潜在蓝色碳汇[J]. 中国科学院院刊, 36(3): 270-278.
|
| [5] |
孙云明, 宋金明, 2002. 中国海洋碳循环生物地球化学过程研究的主要进展(1998-2002)[J]. 海洋科学进展, 20(3): 110-118.
|
| [6] |
肖应凯, 张艳灵, 刘卫国, 等, 2012. 造礁珊瑚δ18Ocarb与海水表面盐度的相关性研究-不同盐度下的珊瑚养殖实验[J]. 地球环境学报, 3(4): 969-981.
|
| [7] |
严宏强, 余克服, 谭烨辉, 2009. 珊瑚礁区碳循环研究进展[J]. 生态学报, 29(11): 6207-6215.
|
| [8] |
张云, 2008. 夏季渤海海—气界面CO2通量及主要影响机制分析[D]. 青岛: 中国海洋大学: 4-10.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}