
海南临高红牌—马袅沿岸海域造礁石珊瑚群落结构及其环境影响因子
|
罗勇(1991—), 男, 贵州省遵义市人, 助理研究员, 研究方向为珊瑚礁生态学。email: |
Copy editor: 林强
收稿日期: 2023-06-15
修回日期: 2023-07-22
网络出版日期: 2023-08-15
基金资助
国家自然科学基金项目(41976120)
国家自然科学基金项目(42276124)
国家重点研发计划项目(2021YFC31005001)
海南省自然科学基金项目(423MS130)
Community structure of reef-building corals and their environmental impact factors in the coastal waters of Hongpai-Maniao, Lingao, Hainan
Copy editor: LIN Qiang
Received date: 2023-06-15
Revised date: 2023-07-22
Online published: 2023-08-15
Supported by
National Natural Science Foundation of China(41976120)
National Natural Science Foundation of China(42276124)
National Key Research and Development Program(2021YFC31005001)
Natural Science Foundation of Hainan Province(423MS130)
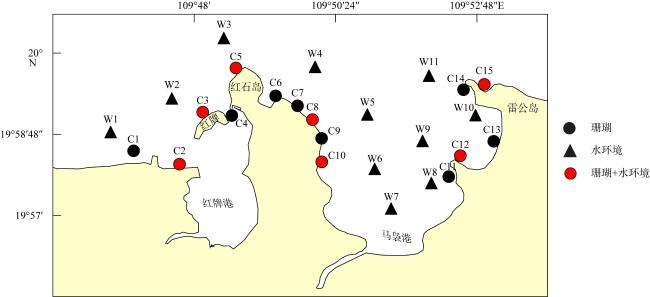
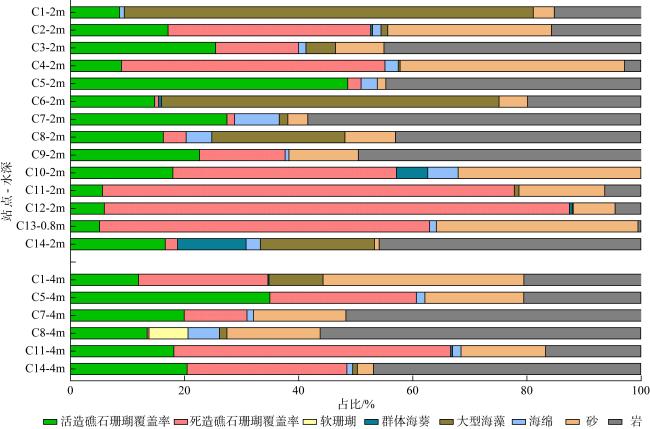
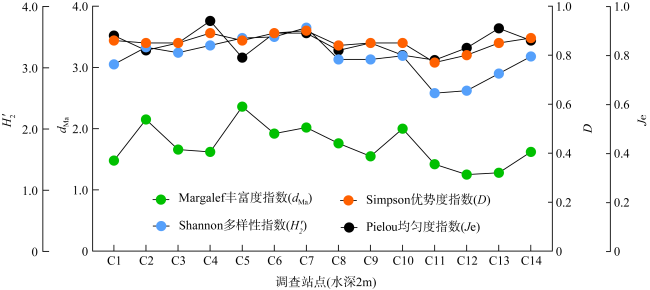
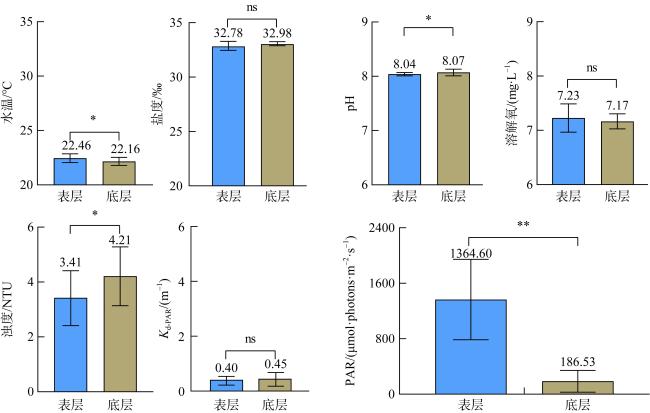
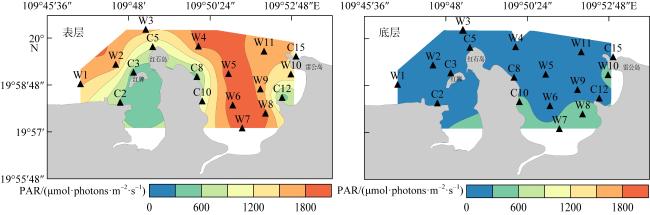
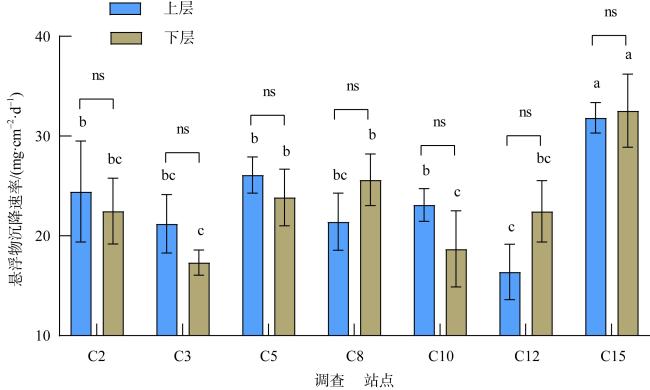
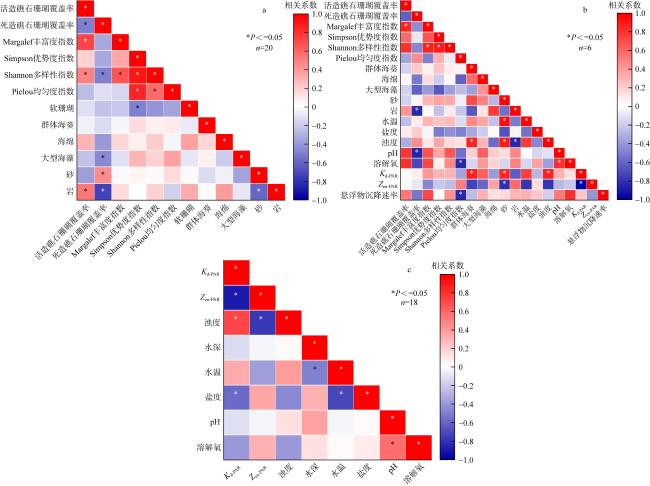
为查明海南临高红牌—马袅沿岸海域造礁石珊瑚群落结构及其环境因子, 于2023年3月18—23日对该区域开展造礁石珊瑚群落、底质类型、悬浮物沉降速率和水环境因子调查。结果显示: (1)本区域造礁石珊瑚共鉴定13科24属42种和7未定种, 其覆盖率均值为(18.05±10.53)%, 集中分布在水深1.0~2.5m; (2)造礁石珊瑚优势种以团块状或皮壳状生长型为主, 死造礁石珊瑚覆盖率(>2年)与活造礁石珊瑚覆盖率、Shannon多样性指数呈显著负相关, 而Shannon多样性指数与Simpson优势度指数和Pielou均匀度指数呈显著正相关, 这表明伴随造礁石珊瑚的死亡, 其覆盖率和物种多样性显著降低; (3)死造礁石珊瑚覆盖率与砂质底覆盖率呈显著正相关, 而与pH呈显著负相关; Pielou均匀度指数与溶解氧和悬浮物沉积物速率呈显著负相关性; 水体浊度与光合有效辐射漫衰减系数Kd-PAR (diffuse attenuation coefficient of the photosynthetically active radiation)呈显著正相关, 且其均值(3.81±1.10)NTU(nephelometric turbidity unit)已超过影响造礁石珊瑚生长的胁迫阈值(3NTU)。这些结果表明, 海底砂质覆盖率、悬浮物沉降速率、浊度、pH、溶解氧和Kd-PAR等是影响该海域造礁石珊瑚群落结构的重要环境因子。综上, 临高红牌—马袅沿岸海域是造礁石珊瑚分布热点之一, 具有较高物种多样性和覆盖率。然而, 该区域造礁石珊瑚垂直分布范围较窄, 且正面临多重环境因子胁迫, 需要采取有效措施进行及时保护。
罗勇 , 黄林韬 , 杨剑辉 , 练健生 , 刘骋跃 , 江雷 , 梁宇娴 , 陈伦举 , 雷新明 , 刘胜 , 黄晖 . 海南临高红牌—马袅沿岸海域造礁石珊瑚群落结构及其环境影响因子[J]. 热带海洋学报, 2024 , 43(3) : 72 -86 . DOI: 10.11978/2023081
To identify the community structure of reef-building corals and their environmental factors in the coastal waters of Hongpai-Maniao, Lingao, Hainan, a survey on the reef-building coral communities, substrate types, sediment accumulation rates and water environmental factors was conducted in the region from 18th to 23th March, 2023. The results showed that: 1) reef-building corals in the region were identified with 42 species in 24 genera of 13 families and 7 undetermined species, with a mean cover of (18.05±10.53)%, concentrated at water depths of 1.0~2.5m; 2) the dominant species of reef-building corals were predominantly massive or encrusting growth types, and dead reef-building coral cover (> 2 years old) was significantly negatively correlated with live reef-building coral cover and Shannon's diversity index, while Shannon's diversity index was significantly positively correlated with Simpson's dominance index and Pielou's evenness index, indicating that the death of reef-building corals was accompanied by a significant reduction in their cover and species diversity; 3) dead reef-building coral cover was significantly positively correlated with sand cover and negatively correlated with pH; the Pielou's evenness index was significantly negatively correlated with dissolved oxygen and sediment accumulation rates; water turbidity was significantly positively correlated with the diffuse attenuation coefficient of the photosynthetically active radiation (Kd-PAR) and its mean value (3.81±1.10) NTU (nephelometric turbidity unit) already exceeded the stress threshold of 3NTU affecting the growth of reef-building corals. These results demonstrate that benthic sand cover, sedimentation rate, turbidity, pH, dissolved oxygen and Kd-PAR are important environmental factors affecting the structure of reef-building coral communities in this area. In summary, the coastal waters of Lingao Hongpai-Maniao is one of the hotspots for the distribution of reef-building corals, with high species diversity and cover. However, the vertical distribution of reef-building corals in this region is narrow and is under stress from multiple environmental factors, requiring effective measures for timely protection.
Key words: Lingao; reef-building corals; community structure; environmental factors; impacts
表1 造礁石珊瑚分布范围与优势种Tab. 1 Distribution range and dominant species of reef-building corals |
| 站点 | 分布水深/m | 样带布设水深/m | 活造礁石珊瑚覆盖率/% | 造礁石珊瑚物种数 | 优势种 | 优势种占活珊瑚覆盖率比例/% | 生长类型 |
|---|---|---|---|---|---|---|---|
| C1 | 1.0~5.0 | 2 | 8.67 | 11 | 澄黄滨珊瑚 Porites lutea | 26.92 | 团块状(massive) |
| 4 | 12.00 | 13 | 斯氏伯孔珊瑚 Bernardpora stutchburyi | 26.39 | 皮壳状(encrusting) | ||
| C2 | 0.5~3.5 | 2 | 17.17 | 17 | 斯氏伯孔珊瑚 Bernardpora stutchburyi | 32.04 | 皮壳状(encrusting) |
| C3 | 0.5~3.5 | 2 | 9.00 | 12 | 十字牡丹珊瑚 Pavona decussata | 16.67 | 指状(digitate) |
| C4 | 0.5~3.5 | 2 | 25.50 | 14 | 粗糙腔星珊瑚 Coelastrea aspera | 30.07 | 团块状(massive) |
| C5 | 0.5~6.5 | 2 | 48.67 | 21 | 角孔珊瑚 Goniopora sp. | 30.48 | 皮壳状(encrusting) |
| 4 | 35.00 | 26 | 多孔同星珊瑚 Plesiastrea versipora | 33.81 | 团块状(massive) | ||
| C6 | 1.5~7.0 | 2 | 14.83 | 15 | 斯托科斯角孔珊瑚 Goniopora stokesi | 20.22 | 柱状(columnar) |
| C7 | 0.5~9.0 | 2 | 27.50 | 17 | 多孔同星珊瑚 Plesiastrea versipora | 17.58 | 团块状(massive) |
| 4 | 20.00 | 18 | 多孔同星珊瑚 Plesiastrea versipora | 37.50 | 团块状(massive) | ||
| C8 | 0.5~5.0 | 2 | 16.33 | 14 | 盘星珊瑚 Dipsastraea sp. | 28.57 | 团块状(massive) |
| 4 | 13.50 | 11 | 多孔同星珊瑚 Plesiastrea versipora | 50.62 | 团块状(massive) | ||
| C9 | 0.5~4.0 | 2 | 22.67 | 13 | 多孔同星珊瑚 Plesiastrea versipora | 28.68 | 团块状(massive) |
| C10 | 0.5~3.0 | 2 | 18.00 | 16 | 大盘星珊瑚 Dipsastraea maxima | 25.00 | 团块状(massive) |
| C11 | 0.5~5.0 | 2 | 5.67 | 10 | 板叶角蜂巢珊瑚 Favites complanata | 38.24 | 团块状(massive) |
| 4 | 18.17 | 12 | 秘密角蜂巢珊瑚 Favites abdita | 54.13 | 团块状(massive) | ||
| C12 | 0.5~3.5 | 2 | 6.00 | 9 | 海洋盘星珊瑚 Dipsastraea maritima | 33.33 | 团块状(massive) |
| C13 | 0.5~6.0 | 2 | 16.67 | 13 | 澄黄滨珊瑚 Porites lutea | 23.00 | 团块状(massive) |
| 4 | 20.50 | 13 | 紫小星珊瑚 Leptastrea purpurea | 33.33 | 皮壳状(encrusting) | ||
| C14 | 0.5~1.5 | 0.8 | 5.17 | 10 | 斯托科斯角孔珊瑚 Goniopora stokesi | 25.81 | 柱状(columnar) |
注: C6站位缺失4m珊瑚样带 |
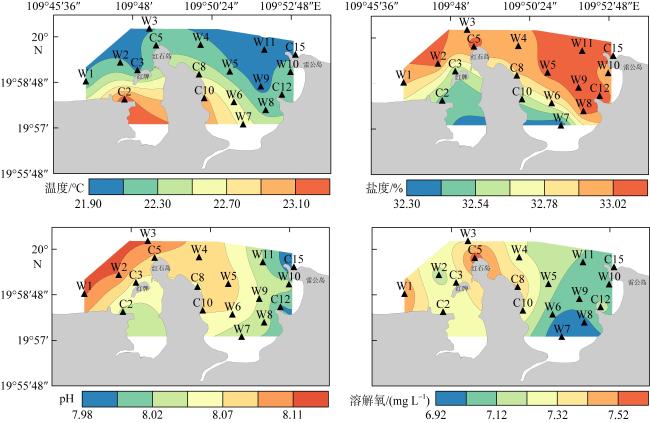
图5 水温、盐度、pH和溶解氧的空间分布Fig. 5 Spatial distribution of temperature, salinity, pH and dissolved oxygen |
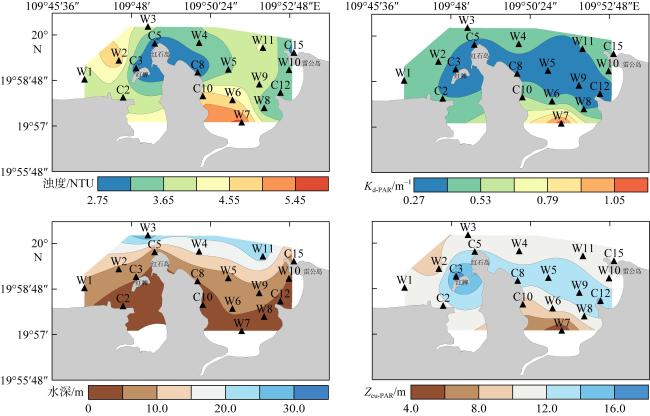
图6 浊度、Kd-PAR、水深和Zeu-PAR的空间分布Fig. 6 Spatial distribution of turbidity, diffuse attenuation coefficient of the photosynthetically active radiation (Kd-PAR), depth and euphotic depth (Zeu-PAR) |
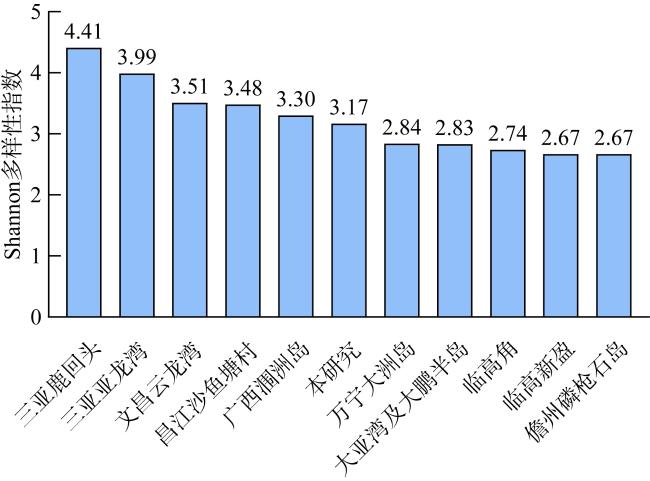
图10 本研究造礁石珊瑚物种多样性与其他区域之间的比较数据来源: 三亚鹿回头(周红英 等, 2017); 三亚亚龙湾(黄丁勇 等, 2020); 文昌云龙湾(周红英 等, 2017); 昌江沙鱼塘村(周红英 等, 2017); 广西涠洲岛(杨振雄 等, 2021); 万宁大洲岛(周红英 等, 2017); 临高角(黄晖 等, 2012); 临高新盈(黄晖 等, 2012); 大亚湾及大鹏半岛(郭峰 等, 2022); 儋州磷枪石岛(周红英 等, 2017) Fig. 10 Comparison of reef-forming coral species diversity between this study and other areas |
表2 临高红牌—马袅沿岸海域造礁石珊瑚物种名录及其与周边海域的对比Tab. 2 List of reef-building coral species in the coastal waters of Lingao’s Hongpai-Maniao and their comparison with surrounding waters |
| 类群 | 科 | 属 | 种 | 临高新盈 和临高角 | 儋州磷 枪石岛Ⅰ | 昌江 海尾 | 儋州磷 枪石岛Ⅱ | 儋州白马井、南华墟和磷枪石岛 | 临高红牌-马袅海域(本研究) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 复杂 类群 | 鹿角 珊瑚科 | 鹿角珊瑚属 | 指形鹿角珊瑚 Acropora digitifera | + | + | |||||||
| 粗野鹿角珊瑚 Acropora humilis | + | + | + | + | ||||||||
| 风信子鹿角珊瑚 Acropora hyacinthus | + | + | ||||||||||
| 多孔鹿角珊瑚 Acropora millepora | + | + | + | |||||||||
| 美丽鹿角珊瑚 Acropora muricata | + | + | ||||||||||
| 鼻形鹿角珊瑚 Acropora nasuta | + | + | ||||||||||
| 霜鹿角珊瑚 Acropora pruinosa | + | + | ||||||||||
| 佳丽鹿角珊瑚 Acropora pulchra | + | + | ||||||||||
| 单独鹿角珊瑚 Acropora solitaryensis | + | |||||||||||
| 鹿角珊瑚 Acropora sp. | + | + | ||||||||||
| 隆起鹿角珊瑚 Acropora tumida | + | |||||||||||
| 壮实鹿角珊瑚 Acropora valida | + | + | ||||||||||
| 星孔珊瑚属 | 多星孔珊瑚 Astreopora myriophthalma | + | ||||||||||
| 蔷薇珊瑚属 | 繁锦蔷薇珊瑚 Montipora efflorescens | + | ||||||||||
| 横错蔷薇珊瑚 Montipora gaimardi | + | |||||||||||
| 弯柔蔷薇珊瑚 Montipora mollis | + | |||||||||||
| 单星蔷薇珊瑚 Montipora monasteriat | + | |||||||||||
| 翼形蔷薇珊瑚 Montipora peltiformis | + | + | + | |||||||||
| 蔷薇珊瑚 Montipora sp. | + | + | + | |||||||||
| 膨胀蔷薇珊瑚 Montipora turgescens | + | + | ||||||||||
| 滨珊瑚科 | 角孔珊瑚属 | 柱形角孔珊瑚 Goniopora columna | + | + | + | |||||||
| 大角孔珊瑚 Goniopora djiboutiensis | + | + | + | |||||||||
| 小角孔珊瑚 Goniopora minor | + | |||||||||||
| 诺福克角孔珊瑚 Goniopora norfolkensis | + | |||||||||||
| 潘朵拉角孔珊瑚 Goniopora pandoraensis | + | |||||||||||
| 扁平角孔珊瑚 Goniopora planulata | + | + | + | + | ||||||||
| 角孔珊瑚 Goniopora sp. | + | + | ||||||||||
| 斯托科斯角孔珊瑚 Goniopora stokesi | + | |||||||||||
| 伯孔珊瑚属 | 斯氏伯孔珊瑚 Bernardpora stutchburyi | + | + | + | + | |||||||
| 滨珊瑚属 | 亚氏滨珊瑚 Porites aranetai | + | ||||||||||
| 团块滨珊瑚 Porites lobata | + | + | ||||||||||
| 澄黄滨珊瑚 Porites lutea | + | + | + | + | + | + | ||||||
| 普哥滨珊瑚 Porites pukoensis | + | + | ||||||||||
| 坚实滨珊瑚 Porites solida | + | |||||||||||
| 滨珊瑚 Porites sp. | + | + | ||||||||||
| 菌珊瑚科 | 厚丝珊瑚属 | 标准厚丝珊瑚 Pachyseris speciosa | + | |||||||||
| 牡丹珊瑚属 | 十字牡丹珊瑚 Pavona decussata | + | + | + | + | + | ||||||
| 叶形牡丹珊瑚 Pavona frondifera | + | |||||||||||
| 真叶珊瑚科 | 真叶珊瑚属 | 联合真叶珊瑚 Euphyllia cristata | + | |||||||||
| 复杂 类群 | 真叶 珊瑚科 | 真叶珊瑚属 | 缨真叶珊瑚 Euphyllia fimbriata | + | ||||||||
| 盔形珊瑚属 | 稀杯盔形珊瑚 Galaxea astreata | + | + | + | ||||||||
| 丛生盔形珊瑚 Galaxea fascicularis | + | + | + | + | + | + | ||||||
| 铁星 珊瑚科 | 假铁星珊瑚属 | 假铁星珊瑚 Pseudosiderastrea tayamai | + | |||||||||
| 木珊瑚科 | 陀螺珊瑚属 | 皱折陀螺珊瑚 Turbinaria mesenterina | + | + | + | |||||||
| 盾形陀螺珊瑚 Turbinaria peltata | + | + | + | + | + | |||||||
| 坚实类群 | 星群 珊瑚科 | 柱群珊瑚属 | 罩柱群珊瑚 Stylocoeniella guentheri | + | + | + | ||||||
| 杯形珊瑚科 | 杯形珊瑚属 | 鹿角杯形珊瑚 Pocillopora damicornis | + | + | ||||||||
| 埃氏杯形珊瑚 Pocillopora eydouxi | + | |||||||||||
| 多曲杯形珊瑚 Pocillopora meandrina | + | |||||||||||
| 疣状杯形珊瑚 Pocillopora verrucosa | + | |||||||||||
| 石芝珊瑚科 | 石芝珊瑚属 | 石芝珊瑚 Fungia fungites | + | |||||||||
| 足柄珊瑚属 | 壳形足柄珊瑚 Podabacia crustacea | + | ||||||||||
| 沙珊瑚科 | 沙珊瑚属 | 深室沙珊瑚 Psammocora contigua | + | + | + | |||||||
| 沙珊瑚 Psammocora sp. | + | |||||||||||
| 筛珊瑚科 | 筛珊瑚属 | 吞蚀筛珊瑚 Coscinaraea exesa | + | + | ||||||||
| 筛珊瑚 Coscinaraea sp. | + | |||||||||||
| 黑星 珊瑚科 | 黑星珊瑚属 | 黑星珊瑚 Oulastrea crispata | + | + | ||||||||
| 叶状 珊瑚科 | 棘星珊瑚属 | 棘星珊瑚 Acanthastrea echinata | + | + | ||||||||
| 联合棘星珊瑚 Acanthastrea hemprichii | + | |||||||||||
| 刺叶珊瑚属 | 粗糙刺叶珊瑚 Echinophyllia aspera | + | ||||||||||
| 叶状珊瑚属 | 菌形叶状珊瑚 Lobophyllia agaricia | + | ||||||||||
| 伞房叶状珊瑚 Lobophyllia corymbosa | + | + | ||||||||||
| 褶曲叶状珊瑚 Lobophyllia flabelliformis | + | |||||||||||
| 赫氏叶状珊瑚 Lobophyllia hemprichii | + | |||||||||||
| 辐射叶状珊瑚 Lobophyllia radians | + | |||||||||||
| 裸肋 珊瑚科 | 刺孔珊瑚属 | 宝石刺孔珊瑚 Echinopora gemmacea | + | |||||||||
| 刺星珊瑚属 | 小叶刺星珊瑚 Cyphastrea microphthalma | + | ||||||||||
| 日本刺星珊瑚 Cyphastrea japonica | + | |||||||||||
| 锯齿刺星珊瑚 Cyphastrea serailia | + | + | + | + | + | + | ||||||
| 菊花珊瑚属 | 粗糙菊花珊瑚 Goniastrea aspera | + | + | + | ||||||||
| 梳状菊花珊瑚 Goniastrea pectinata | + | + | ||||||||||
| 网状菊花珊瑚 Goniastrea retiformis | + | + | ||||||||||
| 菊花珊瑚 Goniastrea sp. | + | + | ||||||||||
| 刺柄珊瑚属 | 腐蚀刺柄珊瑚 Hydnophora exesa | + | + | + | + | + | ||||||
| 圆星珊瑚属 | 曲圆星珊瑚 Astrea curta | + | + | |||||||||
| 腔星珊瑚属 | 粗糙腔星珊瑚 Coelastrea aspera | + | ||||||||||
| 帛琉腔星珊瑚 Coelastrea palauensis | + | + | ||||||||||
| 盘星珊瑚属 | 黄癣盘星珊瑚 Dipsastraea favus | + | + | |||||||||
| 蜥岛盘星珊瑚 Dipsastraea lizardensis | + | |||||||||||
| 海洋盘星珊瑚 Dipsastraea maritima | + | + | ||||||||||
| 大盘星珊瑚 Dipsastraea maxima | + | |||||||||||
| 罗图马盘星珊瑚 Dipsastraea rotumana | + | + | + | + | ||||||||
| 标准盘星珊瑚 Dipsastraea speciosa | + | + | + | + | + | |||||||
| 坚实类群 | 裸肋 珊瑚科 | 盘星珊瑚属 | 盘星珊瑚 Dipsastraea sp. | + | ||||||||
| 扁脑珊瑚属 | 尖边扁脑珊瑚 Platygyra acuta | + | ||||||||||
| 肉质扁脑珊瑚 Platygyra carnosa | + | + | + | |||||||||
| 交替扁脑珊瑚 Platygyra crosslandi | + | + | + | |||||||||
| 精巧扁脑珊瑚 Platygyra daedalea | + | + | + | |||||||||
| 小扁脑珊瑚 Platygyra pini | + | |||||||||||
| 中华扁脑珊瑚 Platygyra sinensis | + | |||||||||||
| 扁脑珊瑚 Platygyra sp. | + | + | ||||||||||
| 琉球扁脑珊瑚 Platygyra ryukyuensis | + | |||||||||||
| 八重山扁脑珊瑚 Platygyra yaeyamaensis | + | |||||||||||
| 角蜂巢珊瑚属 | 秘密角蜂巢珊瑚 Favites abdita | + | + | + | + | + | ||||||
| 尖丘角蜂巢珊瑚 Favites acuticollis | + | |||||||||||
| 中华角蜂巢珊瑚 Favites chinensis | + | + | + | |||||||||
| 板叶角蜂巢珊瑚 Favites complanata | + | + | ||||||||||
| 多弯角蜂巢珊瑚 Favites flexuosa | + | + | + | |||||||||
| 小五边角蜂巢珊瑚 Favites micropentagonus | + | |||||||||||
| 五边角蜂巢珊瑚 Favites pentagona | + | + | + | + | ||||||||
| 角蜂巢珊瑚 Favites sp. | + | + | + | |||||||||
| 同星珊瑚科 | 同星珊瑚属 | 多孔同星珊瑚 Plesiastrea versipora | + | + | + | |||||||
| 未定类群 | 未定 | 小星珊瑚属 | 白斑小星珊瑚 Leptastrea pruinosa | + | ||||||||
| 紫小星珊瑚 Leptastrea purpurea | + | + | ||||||||||
| 总计 | 15 | 33 | 104 | 19 | 45 | 29 | 19 | 55 | 49 | |||
注: 临高新盈和临高角、儋州磷枪石岛Ⅰ、昌江海尾、儋州磷枪石岛Ⅱ及儋州白马井、南华墟和磷枪石岛的珊瑚种类数据分别来源于黄晖 等(2012)、黄晖 等(2012)、牛文涛 等 (2010)、周红英 等 (2017)及廖宝林 等(2020) |
| [1] |
高勤峰, 张恭, 董双林, 2019. 网箱养殖生态学研究进展[J]. 中国海洋大学学报, 49(3): 7-17.
|
| [2] |
郭峰, 肖家光, 田鹏, 等, 2022. 大亚湾及大鹏半岛沿岸造礁石珊瑚现状与生态脆弱性评价[J]. 应用海洋学学报, 41(4): 568-582.
|
| [3] |
胡文佳, 张典, 廖宝林, 等, 2021. 中国大陆沿岸造礁石珊瑚适生区及保护空缺分析[J]. 中国环境科学, 41(1): 401-411.
|
| [4] |
黄丁勇, 李元超, 王建佳, 等, 2020. 亚龙湾西岸造礁石珊瑚种类多样性及恢复潜力分析[J]. 海洋与湖沼, 51(3): 444-455.
|
| [5] |
黄晖, 尤丰, 练健生, 等, 2012. 海南岛西北部海域珊瑚礁造礁石珊瑚种类组成与分布[J]. 海洋科学, 36(9): 64-74.
|
| [6] |
黄建中, 魏宇衡, 顾志峰, 等, 2020. 海南西岛珊瑚群落变化及其影响因素[J]. 热带海洋学报, 39(6): 103-113.
|
| [7] |
黄林韬, 黄晖, 江雷, 2020. 中国造礁石珊瑚分类厘定[J]. 生物多样性, 28(4): 515-523.
|
| [8] |
孔凡洲, 于仁成, 徐子钧, 等, 2012. 应用Excel软件计算生物多样性指数[J]. 海洋科学, 36(4): 57-62.
|
| [9] |
李长青, 夏利栋, 沈贝祺, 等, 2022. 海南三亚珊瑚礁国家级自然保护区珊瑚群落近年的动态变化[J]. 海南热带海洋学院学报, 29(5): 61-72.
|
| [10] |
廖宝林, 肖宝华, 覃业曼, 等, 2020. 海南儋州海域造礁石珊瑚种类组成及动态变化研究[J]. 海洋开发与管理, 37(7): 55-61.
|
| [11] |
刘丽, 李泽鹏, 申玉春, 等, 2013. 四种环境因子对澄黄滨珊瑚和斯氏角孔珊瑚胁迫作用研究[J]. 热带海洋学报, 32(3): 72-77.
|
| [12] |
刘苗苗, 沈建伟, 王月, 等, 2011. 雷州半岛徐闻西岸珊瑚岸礁造礁珊瑚群落结构及其演变[J]. 海洋地质与第四纪地质, 31(6): 37-45.
|
| [13] |
牛文涛, 张潇娴, 林荣澄, 等, 2010. 海南昌江沿岸海域石珊瑚的物种多样性及其分布[J]. 台湾海峡, 29(3): 389-393.
|
| [14] |
施祺, 严宏强, 张会领, 等, 2011. 西沙群岛永兴岛礁坡石珊瑚覆盖率的空间变化[J]. 热带海洋学报, 30(2): 10-17.
|
| [15] |
王道儒, 王华接, 李元超, 等, 2011. 雷州半岛珊瑚幼虫补充来源初步研究[J]. 热带海洋学报, 30(2): 26-32.
|
| [16] |
吴川良, 李长青, 张文勇, 等, 2019. 三亚国家级珊瑚礁自然保护区珊瑚礁资源的多样性[J]. 热带生物学报, 10(1): 14-21.
|
| [17] |
吴钟解, 陈石泉, 陈敏, 等, 2013. 海南岛造礁石珊瑚资源初步调查与分析[J]. 海洋湖沼通报, 35(2): 44-50.
|
| [18] |
闫玉科, 魏紫珊, 林志豪, 2021. 海南省深水网箱养殖业发展研究——以临高县为例[J]. 渔业信息与战略, 36(1): 9-15.
|
| [19] |
杨振雄, 张敬怀, 吕向立, 等, 2021. 涠洲岛造礁石珊瑚群落变化特征及其环境影响因子[J]. 生态学报, 41(18): 7168-7179.
|
| [20] |
周红英, 姚雪梅, 黎李, 等, 2017. 海南岛周边海域造礁石珊瑚的群落结构及其分布[J]. 生物多样性, 25(10): 1123-1130.
|
| [21] |
周洁, 施祺, 余克服, 2014. 三亚造礁石珊瑚虫黄藻光合作用效率的日周期及其调控因素[J]. 热带海洋学报, 33(1): 81-89.
|
| [22] |
邹仁林, 2001. 中国动物志: 腔肠动物门. 珊瑚虫纲. 石珊瑚目: 造礁石珊瑚[M]. 北京: 科学出版社: 1-242 (in Chinese).
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}