
珊瑚礁生态系统固碳过程及储碳机制研究进展*
*感谢珊瑚生物学和珊瑚礁生态学学科组全体人员对本论文的完成提供的帮助。
|
黄晖(1969—), 女, 江西省分宜县人,研究员, 从事珊瑚生物学与珊瑚礁生态学等方面研究。email: |
Copy editor: 林强
收稿日期: 2023-04-14
修回日期: 2023-07-05
网络出版日期: 2023-08-17
基金资助
海南省重点研发计划项目(ZDYF2023SHFZ131)
国家重点研发计划项目(2021YFF0502800)
Carbon sequestration process and carbon storage mechanism of reef ecosystem in South China Sea*
Copy editor: LIN Qiang
Received date: 2023-04-14
Revised date: 2023-07-05
Online published: 2023-08-17
Supported by
Key Research and Development Project of Hainan Province, China(ZDYF2023SHFZ131)
National Key Research and Development Program of China(2021YFF0502800)
黄晖 , 袁翔城 , 宋严 , 李颖心 , 周伟华 , 龙爱民 . 珊瑚礁生态系统固碳过程及储碳机制研究进展*[J]. 热带海洋学报, 2024 , 43(3) : 13 -21 . DOI: 10.11978/2023049
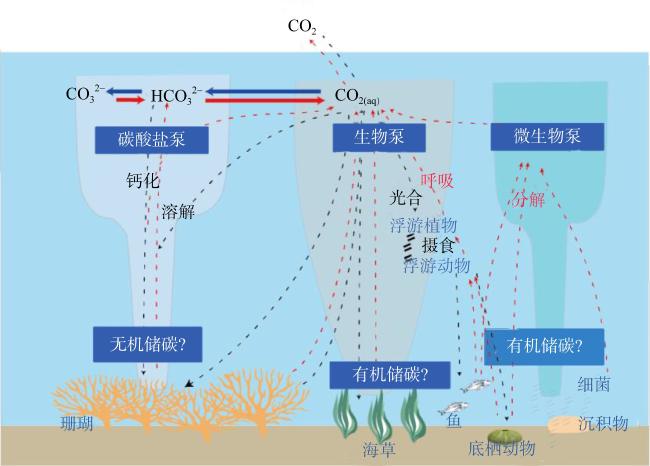
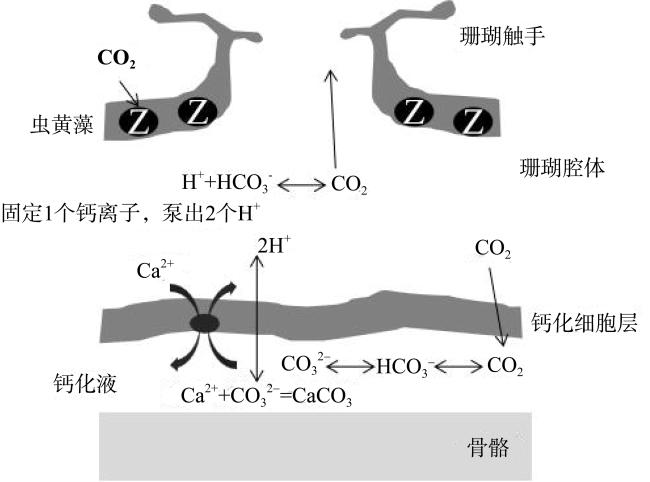
Coral reef ecosystems play a crucial role in global carbon cycling and climate change. They serve as potential blue carbon sinks, with coral reefs in China covering approximately 11% of the global area. The annual carbon sequestration of these reefs is estimated to be 3.5×106~4.5×106 t. The total carbon storage capacity of coral reef ecosystems is immense, and they have significant potential value as carbon reservoirs in carbon trading. This article provides a review of carbon sequestration processes and storage mechanisms in coral reef ecosystems, including carbonate pumping, biological pumping, and microbially-mediated carbon pumping. The research progress on these three carbon pumps is discussed. Additionally, the article highlights the limitations and challenges in current research and proposes future research directions, aiming to provide scientific basis for the conservation and management of coral reef ecosystems in the South China Sea.
| [1] |
蔡榕硕, 郭海峡,
|
| [2] |
杜明卉, 李昌达, 杨华蕾, 等, 2023. 海岸带蓝碳生态系统碳库规模与投融资机制[J]. 海洋环境科学, 42(2): 294-301.
|
| [3] |
黄晖, 张浴阳, 黄洁英, 等, 2014. 不同移植方法对多种珊瑚移植效果的影响[C]// 2014水域生态环境修复学术研讨会论文集, 2014, 54-63,
|
| [4] |
黄晖, 张浴阳, 刘骋跃, 2020. 热带岛礁型海洋牧场中珊瑚礁生境与资源的修复[J]. 科技促进发展, 16(2): 225-230.
|
| [5] |
焦念志, 刘纪化, 石拓, 等, 2021. 实施海洋负排放践行碳中和战略[J]. 中国科学: 地球科学, 51(4): 632-643.
|
| [6] |
焦念志, 戴民汉, 翦知湣, 等, 2022. 海洋储碳机制及相关生物地球化学过程研究策略[J]. 科学通报, 67(15): 1600-1606.
|
| [7] |
李银强, 2022. 西沙群岛珊瑚藻记录之近2000万年以来的海平面变化和珊瑚礁演化过程[D]. 南宁: 广西大学.
|
| [8] |
施祺, 赵美霞, 张乔民, 等, 2009. 海南三亚鹿回头造礁石珊瑚碳酸盐生产力的估算[J]. 科学通报, 54(10): 1471-1479.
|
| [9] |
石拓, 郑新庆, 张涵, 等, 2021. 珊瑚礁: 减缓气候变化的潜在蓝色碳汇[J]. 中国科学院院刊, 36(3): 270-278.
|
| [10] |
唐剑武, 叶属峰, 陈雪初, 等, 2018. 海岸带蓝碳的科学概念、研究方法以及在生态恢复中的应用[J]. 中国科学: 地球科学, 48(6): 661-670.
|
| [11] |
王法明, 唐剑武, 叶思源, 等, 2021. 中国滨海湿地的蓝色碳汇功能及碳中和对策[J]. 中国科学院院刊, 36(3): 241-251.
|
| [12] |
张军晓, 2022. 南海典型珊瑚礁区碳酸盐体系与钙化率研究[D]. 广州: 中国科学院研究生院(南海海洋研究所).
|
| [13] |
张浴阳, 刘骋跃, 王丰国, 等, 2021. 典型近岸退化珊瑚礁的成功修复案例——蜈支洲珊瑚覆盖率的恢复[J]. 应用海洋学学报, 40(1): 26-33.
|
| [14] |
郑新庆, 张涵, 陈彬, 等, 2021. 珊瑚礁生态修复效果评价指标体系研究进展[J]. 应用海洋学学报, 40(1): 126-141.
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
IPCC, 2019. Summary for Policymakers[R/OL]. [2023-04-14]. https://www.ipcc.ch/site/assets/uploads/sites/4/2022/11/SRCCL_SPM.pdf
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
VAN DER HEIJDEN L H,
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}