
基于高通量测序技术的深圳湾真核浮游植物群落结构研究
|
黄圆(1999—), 女, 广东省河源市人, 硕士研究生, 从事藻类分类学研究。email: |
Copy editor: 殷波
收稿日期: 2023-07-03
修回日期: 2023-08-17
网络出版日期: 2023-08-28
基金资助
北京市自然科学基金项目(8232026)
国家自然科学基金项目(41906112)
基础资源调查专项(2018FY100200)
Study on the community structure of eukaryotic phytoplankton in the Shenzhen Bay based on high-throughput sequencing technology
Copy editor: YIN Bo
Received date: 2023-07-03
Revised date: 2023-08-17
Online published: 2023-08-28
Supported by
Beijing Natural Science Foundation(8232026)
National Natural Science Foundation of China(41906112)
National Science and Technology Fundamental Resources Investigation Programme(2018FY100200)
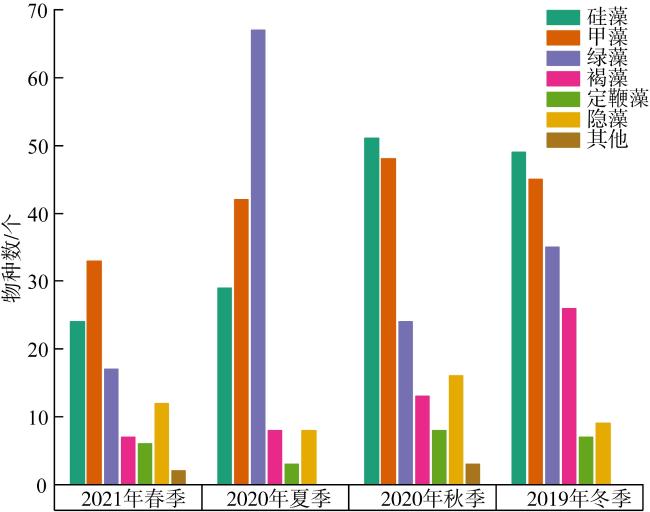
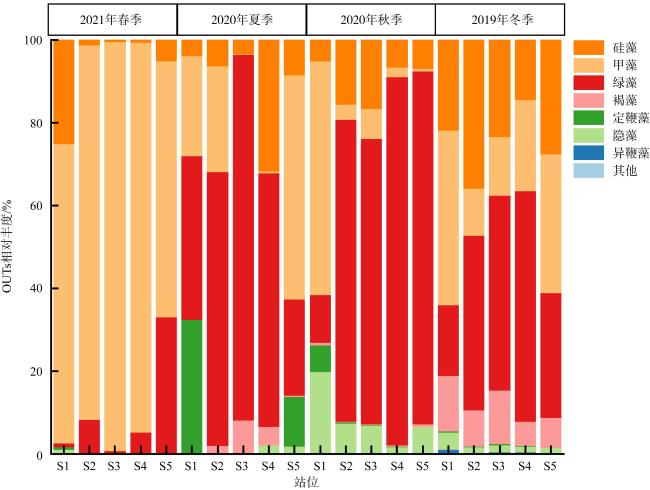
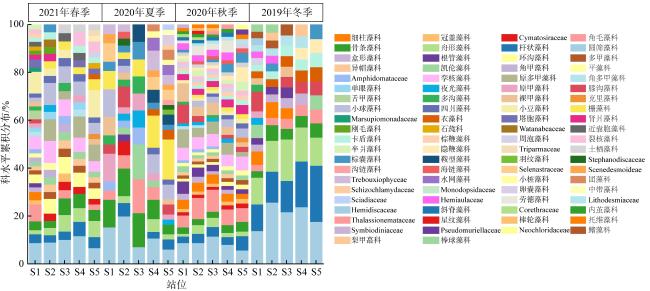
以18S rDNA V4区为目标基因, 采用Illumina Miseq高通量测序技术, 对深圳湾2019年冬季—2021年春季4个季节海水中浮游植物群落结构进行研究, 并整理该海域有害藻华生物的历史纪录数据, 以探讨深圳湾有毒有害浮游植物的群落组成及演替规律。高通量测序结果显示, 深圳湾海域真核浮游植物含硅藻、甲藻、绿藻、褐藻、定鞭藻、隐藻和红藻7个类群, 共82科269种。其中以微微型浮游植物为主要类群, 春季和秋季以微拟球藻(Nannochloris sp.)为优势种, 夏季以球等鞭金藻(Isochrysis galbana)为优势种, 冬季以极小海链藻(Thalassiosira minima)为优势种。冬季浮游植物多样性指数为全年最高。此次调查共检测出有毒有害浮游植物32种, 其中甲藻15种、硅藻13种、褐藻3种、定鞭藻1种。与历史资料相比, 深圳湾海域的有毒有害浮游植物呈微型化和多样性逐渐增加的趋势, 呈现由甲藻或硅藻为主的有毒有害单类群群落演替到以甲藻和硅藻为主的有毒有害双类群群落的趋势。
黄圆 , 岑竞仪 , 梁芊艳 , 吕颂辉 , 王建艳 . 基于高通量测序技术的深圳湾真核浮游植物群落结构研究[J]. 热带海洋学报, 2024 , 43(2) : 21 -33 . DOI: 10.11978/2023093
Using the 18S rDNA V4 region as the target gene, Illumina Miseq high-throughput sequencing technology was applied to study the phytoplankton community in the seawater of the Shenzhen Bay during the four seasons from the winter of 2019 to the spring of 2021, and to compare with the historical data of harmful algal blooms species in this area, in order to explore the community composition and succession pattern of toxic and harmful phytoplankton in the Shenzhen Bay. The high-throughput sequencing results showed that the eukaryotic phytoplankton in the sea waters of the Shenzhen Bay consisted of 7 groups, including Bacillariophyta, Dinoflagellata, Chlorophyta, Ochrophyta, Haptophyta, Cryptophyta, and Rhodophyta, with a total of 82 families and 269 species. Among them, microphytoplankton was the main group, Nannochloris sp. was the dominant species in spring and autumn, Isochrysis galbana was the dominant species in summer, and Thalassiosira minima was the dominant species in winter. The phytoplankton diversity index in winter was the highest in the four seasons. A total of 32 species of toxic and harmful phytoplankton were detected in this survey, including 15 dinoflagellata species, 13 bacillariophyta species, 3 ochrophyta species, and 1 of haptophyta species. Compared with the historical data, the toxic and harmful phytoplankton in the Shenzhen Bay showed a miniaturization trend and an increasing diversity. The monophasic harmful blooms dominated by dinoflagellates or diatoms have been succeeded to polyphasic blooms of dinoflagellates and diatoms.
表2 深圳湾4个季节OTUs注释结果Tab. 2 The amount of annotation OTUs of the four season seawater samples in the Shenzhen Bay |
| 季节 | 序列数 | OTUs总数 | 浮游植物OTUs数目 | 浮游植物OTUs占比/% |
|---|---|---|---|---|
| 2021年春季 | 41964~71230 | 756 | 244 | 32.28 |
| 2020年夏季 | 54698~118350 | 2522 | 660 | 26.17 |
| 2020年秋季 | 69035~73509 | 1058 | 356 | 33.65 |
| 2019年冬季 | 50352~53527 | 2057 | 513 | 24.94 |
表3 深圳湾4个季节浮游植物优势种Tab. 3 Dominant phytoplankton in Shenzhen Bay |
| 季节 | 物种 | 拉丁名 | 优势度 | 出现频率/% | 平均丰度/(×103个·m-3) |
|---|---|---|---|---|---|
| 2021年春季 | 微拟球藻 | Nannochloris sp. | 0.54 | 100 | 43.12 |
| 小环藻 | Cyclotella choctawhatcheeana | 0.17 | 100 | 13.77 | |
| Picochlorum maculatum | Picochlorum maculatum | 0.08 | 80 | 8.13 | |
| 环沟藻 | Gyrodinium sp. | 0.08 | 100 | 6.40 | |
| 新月筒柱藻 | Cylindrotheca closterium | 0.02 | 80 | 2.41 | |
| 2020年夏季 | 球等鞭金藻 | Isochrysis galbana | 0.47 | 100 | 46.71 |
| 小球藻 | Chlorella sorokiniana | 0.07 | 60 | 11.78 | |
| 威氏海链藻 | Thalassiosira weissflogii | 0.08 | 80 | 10.34 | |
| 吉思纳海链藻 | Thalassiosira gessneri | 0.06 | 60 | 9.20 | |
| 双眉藻 | Sellaphora sp. | 0.04 | 60 | 5.91 | |
| 2020年秋季 | 微拟球藻 | Nannochloris sp. | 0.71 | 100 | 103.40 |
| 垂裂莱万藻 | Levanderina fissa | 0.02 | 100 | 2.94 | |
| 骨条藻 | Skeletonema sp. | 0.05 | 100 | 7.61 | |
| 异帽藻 | Heterocapsa sp. | 0.03 | 100 | 3.59 | |
| 海链藻 | Thalassiosira sp. | 0.04 | 100 | 5.63 | |
| 小环藻 | Cyclotella choctawhatcheeana | 0.02 | 100 | 3.10 | |
| 2019年冬季 | 赤潮异弯藻 | Heterosigma akashiwo | 0.26 | 100 | 7.75 |
| 舟形藻 | Navicula sp. | 0.04 | 100 | 1.18 | |
| 亚当斯骨条藻 | Skeletonema ardens | 0.06 | 100 | 1.93 | |
| 菱形藻 | Nitzschia sp. | 0.03 | 100 | 0.88 | |
| 新月筒柱藻 | Cylindrotheca closterium | 0.06 | 100 | 1.79 | |
| 极小海链藻 | Thalassiosira minima | 0.43 | 100 | 12.83 | |
| 梅尼小环藻 | Cyclotella meneghiniana | 0.03 | 80 | 0.93 |
表4 深圳湾浮游植物多样性指数Tab. 4 The diversity indices of phytoplankton in the Shenzhen Bay |
| 季节 | 站位 | Shannon-Wiener多样性指数 | Margalef丰富度指数 | Pielou均匀度指数 |
|---|---|---|---|---|
| 2021年春季 | S1 | 1.03 | 6.03 | 0.25 |
| S2 | 1.20 | 3.37 | 0.35 | |
| S3 | 1.88 | 3.58 | 0.57 | |
| S4 | 1.51 | 3.20 | 0.44 | |
| S5 | 1.11 | 3.83 | 0.30 | |
| 2020年夏季 | S1 | 1.41 | 1.89 | 0.48 |
| S2 | 1.51 | 3.46 | 0.44 | |
| S3 | 1.53 | 1.39 | 0.60 | |
| S4 | 1.25 | 1.94 | 0.41 | |
| S5 | 1.75 | 1.97 | 0.57 | |
| 2020年秋季 | S1 | 2.74 | 8.12 | 0.63 |
| S2 | 1.45 | 7.22 | 0.33 | |
| S3 | 1.44 | 7.02 | 0.33 | |
| S4 | 0.85 | 5.42 | 0.21 | |
| S5 | 0.71 | 4.38 | 0.19 | |
| 2019年冬季 | S1 | 2.96 | 4.57 | 0.84 |
| S2 | 1.38 | 3.06 | 0.41 | |
| S3 | 1.29 | 2.29 | 0.42 | |
| S4 | 1.69 | 2.12 | 0.58 | |
| S5 | 1.93 | 1.66 | 0.71 |
表5 深圳湾4个季节有毒有害浮游植物名录Tab. 5 List of toxic and harmful phytoplankton in the Shenzhen Bay |
| 序号 | 门类 | 种名 | 拉丁文名 | 危害 | 春季 | 夏季 | 秋季 | 冬季 |
|---|---|---|---|---|---|---|---|---|
| 1 | 甲藻门 | 近缘亚历山大藻 | Alexandrium affine | 有毒 | √ | √ | √ | |
| 2 | 广野亚历山大藻 | Alexandrium hiranoi | 有毒 | √ | ||||
| 3 | 链状亚历山大藻 | Alexandrium catenella | 有毒 | √ | ||||
| 4 | 奥斯亚历山大藻 | Alexandrium ostenfeldii | 有毒 | √ | √ | |||
| 5 | 安德森亚历山大藻 | Alexandrium andersonii | 有毒 | √ | ||||
| 6 | 塔玛亚历山大藻 | Alexandrium tamarense | 有毒 | √ | ||||
| 7 | 渐尖鳍藻 | Dinophysis acuminata | 有毒 | √ | ||||
| 8 | 具刺膝沟藻 | Gonyaulax spinifera | 有毒 | √ | √ | √ | ||
| 9 | 垂裂莱万藻(原名条纹环沟藻) | Levanderina fissa | 有害 | √ | ||||
| 10 | 剧毒卡尔藻 | Karlodinium veneficum | 有毒 | √ | √ | |||
| 11 | 夜光藻 | Noctiluca scintillans | 有害 | √ | √ | √ | √ | |
| 12 | 凹面原甲藻 | Prorocentrum concavum | 有毒 | √ | √ | |||
| 13 | 钝齿原甲藻(原名东海原甲藻) | Prorocentrum obtusidens | 有害 | √ | ||||
| 14 | 巴哈马麦甲藻 | Pyrodinium bahamense | 有毒 | √ | ||||
| 15 | 双胞多沟藻 | Polykrikos geminatum | 有害 | √ | ||||
| 16 | 硅藻门 | 薄壁几内亚藻 | Guinardia flaccida | 有害 | √ | |||
| 17 | 柔弱角毛藻 | Chaetoceros debilis | 有害 | √ | ||||
| 18 | 并基角毛藻 | Chaetoceros decipiens | 有害 | √ | ||||
| 19 | 平孢角毛藻 | Chaetoceros laevisporus | 有害 | √ | ||||
| 20 | 格氏圆筛藻 | Coscinodiscus granii | 有害 | √ | ||||
| 21 | 琼氏圆筛藻 | Coscinodiscus jonesianus | 有害 | √ | √ | |||
| 22 | 辐射圆筛藻 | Coscinodiscus radiatus | 有害 | √ | √ | |||
| 23 | 环纹娄氏藻 | Lauderia annulata | 有害 | √ | ||||
| 24 | 丹麦细柱藻 | Leptocylindrus danicus | 有害 | √ | √ | |||
| 25 | 伦德海链藻 | Thalassiosira lundiana | 有害 | √ | √ | |||
| 26 | 极小海链藻 | Thalassiosira minima | 有害 | √ | √ | |||
| 27 | 威氏海链藻 | Thalassiosira weissflogii | 有害 | √ | ||||
| 28 | 萎软海链藻 | Thalassiosira mala | 有害 | √ | ||||
| 29 | 褐藻门 | 海洋卡盾藻 | Chattonella marina | 有害 | √ | |||
| 30 | 盐生卡盾藻 | Chattonella subsalsa | 有毒 | √ | √ | |||
| 31 | 赤潮异弯藻 | Heterosigma akashiwo | 有害 | √ | √ | √ | ||
| 32 | 定鞭藻门 | 球形棕囊藻 | Phaeocystis globosa | 有害 | √ | √ |
注: √表示此季节出现的有毒有害浮游植物; 空白表示此季节未出现的有毒有害浮游植物 |
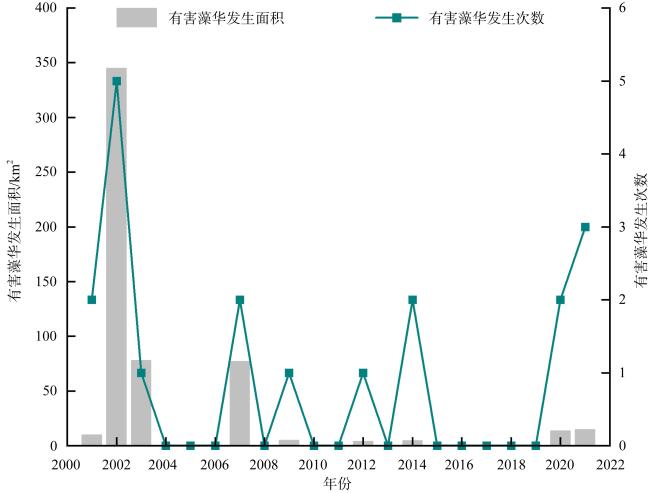
图5 深圳海域有毒有害浮游植物赤潮爆发情况Fig. 5 Toxic and harmful algal blooms in the coastal waters of Shenzhen from 1980 to 2021 |
表6 深圳湾有毒有害浮游植物粒级变化Tab. 6 Changes in the size of toxic and harmful phytoplankton in the Shenzhen Bay |
| 粒径水平 | 物种数目(占比) | ||||
|---|---|---|---|---|---|
| 20世纪80年代 | 20世纪90年代 | 2000—2010年 | 2010—2021年 | 本研究 | |
| 微型浮游植物 | 1 (8.33%) | 4 (16.67%) | 4 (15.38%) | 3 (16.67%) | 7 (21.88%) |
| 小型浮游植物 | 11 (91.67%) | 20 (83.33%) | 22 (84.62%) | 15 (83.33%) | 25 (78.12%) |
| 总数 | 12 | 24 | 26 | 18 | 32 |
| [1] |
陈思, 2020. 基于层次分析法的深圳湾禁渔效果评价研究[D]. 上海:上海海洋大学:60-63.
|
| [2] |
陈思, 陈海刚, 田斐, 等, 2021. 深圳湾浮游植物群落结构特征及其与环境因子的关系[J]. 生态科学, 40(1): 9-16.
|
| [3] |
广东省海洋与渔业局, 广东省海洋环境质量公报[EB/OL]. (2009, 2010). http://gdee.gd.gov.cn/hjzkgb/index_2.html (in Chinese).
|
| [4] |
广东省海洋与渔业局, 广东省海洋环境状况公报[EB/OL]. (2011, 2012, 2013, 2014, 2015, 2016, 2017). http://gdeegd.gov.cn/hjzkgb/index_2.html (in Chinese).
|
| [5] |
广东省生态环境厅. 广东省生态环境状况公报[EB/OL]. (2018, 2019, 2020, 2021). http://gdee.gd.gov.cn/hjzkgb/index_2.html (in Chinese).
|
| [6] |
桓清柳, 庞仁松, 周秋伶, 等, 2016. 深圳近岸海域氮、磷营养盐变化趋势及其与赤潮发生的关系[J]. 海洋环境科学, 35(6): 908-914.
|
| [7] |
冷科明, 江天久, 2004. 深圳海域近20年赤潮发生的特征分析[J]. 生态科学, 23(2): 166-170.
|
| [8] |
李港, 林妙丽, 陈诚, 等, 2021. 京杭运河苏北段秋冬季浮游植物群落结构特征及其与环境因子的关系[J]. 水生态学杂志, 42(5): 119-126.
|
| [9] |
刘华健, 黄良民, 谭烨辉, 等, 2017. 珠江口浮游植物叶绿素a和初级生产力的季节变化及其影响因素[J]. 热带海洋学报, 36(1): 81-91.
|
| [10] |
邱阳凌, 林育青, 刘俊杰, 等, 2018. 淮河干流及主要支流夏季浮游植物群落生物多样性评价[J]. 环境科学学报, 38(4): 1665-1672.
|
| [11] |
孙金水, 戴纪翠, 倪晋仁, 等, 2010a. 深圳湾海域浮游植物的生态特征[J]. 环境科学, (1): 63-68.
|
| [12] |
孙金水,
|
| [13] |
王朝晖, 齐雨藻, 尹伊伟, 等, 2001. 1998年春深圳湾环节环沟藻赤潮及其发生原因的探讨[J]. 海洋科学, 25(5): 47-50.
|
| [14] |
吴振斌, 贺锋, 付贵萍, 等, 2002. 深圳湾浮游生物和底栖动物现状调查研究[J]. 海洋科学, 26(8): 58-64.
|
| [15] |
向晨晖, 刘甲星, 柯志新, 等, 2021. 大亚湾浮游植物粒级结构和种类组成对淡澳河河口水加富的响应[J]. 热带海洋学报, 40(2): 49-60.
|
| [16] |
袁超, 徐宗军, 张学雷, 2015. 2010—2011年深圳湾浮游植物季节变化及其与环境因子关系[J]. 海洋湖沼通报, (1): 112-120.
|
| [17] |
张冬鹏, 黎晓涛, 黄远峰, 等, 2001. 深圳沿海浮游植物组成及赤潮发生趋势分析[J]. 暨南大学学报(自然科学与医学版), 22(5): 122-126.
|
| [18] |
张才学, 周凯, 孙省利, 等, 2010. 深圳湾浮游动物的群落结构及季节变化[J]. 生态环境学报, 19(11): 2686-2692.
|
| [19] |
赵东至, 2010. 中国典型海域赤潮灾害发生规律[M]. 北京: 海洋出版社:269- 273.
|
| [20] |
朱广伟, 施坤, 李未, 等, 2020. 太湖蓝藻水华的年度情势预测方法探讨[J]. 湖泊科学, 32(5): 1421-1431.
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}