
广西涠洲岛蛭态轮虫物种多样性及生境偏好研究
|
陈俊强(1998—), 男, 广东省中山市人, 硕士研究生, 从事轮虫分类和生态学研究。email: |
Copy editor: 殷波
收稿日期: 2023-06-19
修回日期: 2023-08-20
网络出版日期: 2023-09-04
基金资助
国家自然科学基金面上项目(41673080)
广东省基础研究与应用基础研究基金项目(2021A1515010814)
Species diversity and habitat preference of bdelloid rotifers in the Weizhou island, Guangxi
Copy editor: YIN Bo
Received date: 2023-06-19
Revised date: 2023-08-20
Online published: 2023-09-04
Supported by
National Natural Science Foundation of China(41673080)
Guangdong Basic and Applied Basic Research Foundation(2021A1515010814)
岛屿生境和生物群落组成独特, 但有关岛屿蛭态轮虫的研究报道较少。为研究岛屿环境蛭态轮虫的物种多样性, 于2021—2022年在广西涠洲岛苔藓、土壤、竹叶、松树落叶、其他落叶、带柄植物花瓣和燃烧后叶杆共7种生境进行调查, 共记录蛭态轮虫3科8属53种, 其中中国新记录种3种, 结果显示涠洲岛蛭态轮虫物种多样性高(占全球已报道蛭态轮虫物种数量10.6%)。前4种生境间物种丰富度差异显著(p<0.0001), 且不同生境间物种组成差异性显著大于同类生境内物种组成差异(p=0.001), 表明蛭态轮虫生境偏好明显; 指示种分析(IndVal)表明, 共有10种蛭态轮虫显著偏好某一生境。
陈俊强 , 汪文博 , 王庆 , 杨宇峰 . 广西涠洲岛蛭态轮虫物种多样性及生境偏好研究[J]. 热带海洋学报, 2024 , 43(2) : 81 -91 . DOI: 10.11978/2023083
The island habitat and biological community composition are unique, but there are few reports on the island bdelloid rotifers. During 2021—2022, we investigated seven different habitats, i.e., moss, soil, leaf litters of bamboo, pine and other, petiolate plant petals and burned leaf stems, in the Weizhou island, Guangxi, to investigate species diversity and habitat preference of bdelloid rotifers. In the Weizhou island, a total of 53 species of bdelloid rotifers in three families, eight genera and 53 species were recorded, of which three species were newly found in China. The results showed that the species diversity of bdelloid rotifers in the island was high (accounting for 10.6% of the global reported bdelloid rotifer species). There was significant difference in species richness among the first four habitats (p < 0.0001), and the difference of species composition between different habitats is significantly greater than that within the same habitat (p = 0.001). The habitat preference of bdelloid rotifer is obvious. In addition, IndVal index analysis showed that 10 rotifer species had significant preference for special habitats. In the future, we should strengthen the investigation of island habitats, especially rare habitats, for the purpose of finding more species of bdelloid rotifers and enriching their geographical distribution in China.
Key words: bdelloid rotifer; island; habitat preference; new record species; species diversity
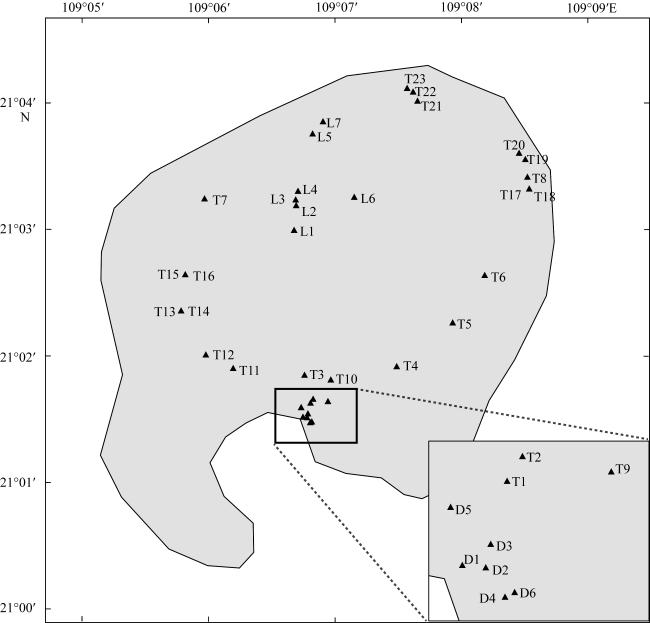
图1 涠洲岛采样位点图该图基于自然资源部标准地图服务网站下载的审图号为GS(2022)4309号标准地图制作, 底图无修改 Fig. 1 Sampling sites in the Weizhou island |
表1 涠洲岛采样点信息Tab. 1 The sampling sites in the Weizhou island |
| 采样点编号 | 采样日期 | 生境类型 | 位置坐标 |
|---|---|---|---|
| D1 | 2021.12.25 | 苔藓 | 21°01′31.18″N, 109°06′44.85″E |
| D2 | 2021.12.25 | 苔藓 | 21°01′30.99″N, 109°06′46.72″E |
| D3 | 2021.12.25 | 苔藓 | 21°01′32.86″N, 109°06′47.11″E |
| D4 | 2021.12.25 | 竹叶 | 21°01′28.66″N, 109°06′48.23″E |
| D5 | 2021.12.25 | 带柄植物花瓣 | 21°01′35.78″N, 109°06′43.92″E |
| D6 | 2021.12.25 | 其他落叶 | 21°01′29.04″N, 109°06′49.01″E |
| T1 | 2022.05.20 | 松树落叶 | 21°01′37.85″N, 109°06′48.41″E |
| T2 | 2022.05.20 | 松树落叶 | 21°01′39.81″N, 109°06′49.63″E |
| T3 | 2022.05.20 | 松树落叶 | 21°01′51.10″N, 109°06′45.49″E |
| T4 | 2022.05.20 | 松树落叶 | 21°01′55.18″N, 109°07′29.31″E |
| T5 | 2022.05.20 | 松树落叶 | 21°02′15.92″N, 109°07′55.89″E |
| T6 | 2022.05.20 | 松树落叶 | 21°02′38.42″N, 109°08′10.98″E |
| T7 | 2022.05.20 | 松树落叶 | 21°03′14.81″N, 109°05′58.09″E |
| T8 | 2022.05.20 | 松树落叶 | 21°03′25.02″N, 109°08′31.24″E |
| T9 | 2022.05.20 | 土壤 | 21°01′38.60″N, 109°06′56.67″E |
| T10 | 2022.05.20 | 土壤 | 21°01′48.87″N, 109°06′58.02″E |
| T11 | 2022.05.20 | 土壤 | 21°01′54.35″N, 109°06′11.71″E |
| T12 | 2022.05.20 | 苔藓 | 21°02′00.77″N, 109°05′58.72″E |
| T13 | 2022.05.20 | 土壤 | 21°02′21.57″N, 109°05′46.95″E |
| T14 | 2022.05.20 | 苔藓 | 21°02′21.57″N, 109°05′46.95″E |
| T15 | 2022.05.20 | 苔藓 | 21°02′38.85″N, 109°05′48.94″E |
| T16 | 2022.05.20 | 土壤 | 21°02′38.85″N, 109°05′48.94″E |
| T17 | 2022.05.20 | 土壤 | 21°03′19.43″N, 109°08′32.16″E |
| T18 | 2022.05.20 | 苔藓 | 21°03′19.43″N, 109°08′32.16″E |
| T19 | 2022.05.20 | 土壤 | 21°03′33.47″N, 109°08′30.27″E |
| T20 | 2022.05.20 | 其他落叶 | 21°03′36.33″N, 109°08′27.31″E |
| T21 | 2022.05.20 | 燃烧后的叶杆 | 21°04′01.04″N, 109°07′39.11″E |
| T22 | 2022.05.20 | 其他落叶 | 21°04′05.46″N, 109°07′37.01″E |
| T23 | 2022.05.20 | 土壤 | 21°04′07.17″N, 109°07′34.15″E |
| L1 | 2022.09.01 | 竹叶 | 21°02′59.77″N, 109°06′40.56″E |
| L2 | 2022.09.01 | 竹叶 | 21°03′11.56″N, 109°06′41.59″E |
| L3 | 2022.09.01 | 竹叶 | 21°03′14.23″N, 109°06′41.36″E |
| L4 | 2022.09.01 | 竹叶 | 21°03′18.36″N, 109°06′42.47″E |
| L5 | 2022.09.01 | 竹叶 | 21°03′45.55″N, 109°06′49.39″E |
| L6 | 2022.09.01 | 其他落叶 | 21°03′15.45″N, 109°07′09.17″E |
| L7 | 2022.09.01 | 竹叶 | 21°03′51.35″N, 109°06′54.29″E |
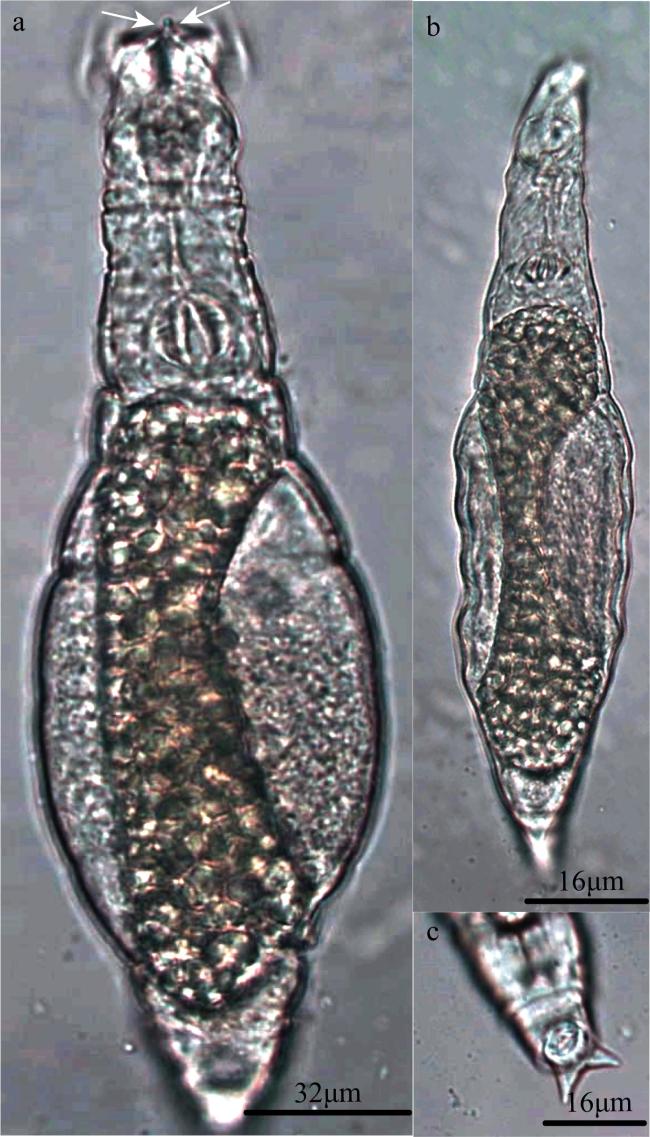
图2 戈氏粗颈轮虫(Bryce, 1912)a. 爬行状态背面观; b, c. 摄食状态背面观(箭头指向舌凸); d, f. 爬行状态足部腹面观(箭头指向趾); e. 摄食状态头部背面观(箭头指向舌凸和上唇片) Fig. 2 Macrotrachela decora (Bryce, 1912). (a) Creeping, dorsal view; (b) and (c) Feeding, dorsal view (an arrow pointing at ligula); (d) and (f) Creeping (foot), ventral view (an arrow pointing at toes); (e) Feeding (head), dorsal view (an arrow pointing at ligula and overlip) |
图4 美丽粗颈轮虫(Murray, 1907)a. 爬行状态背面观(箭头指向褶皱); b, c. 摄食状态背面观(b中箭头指向瓣环和褶皱, c中箭头指向褶皱); d. 摄食状态头部背面观; e. 爬行状态足部背面观; f. 爬行状态足部侧面观; g. 爬行状态足部腹面观 Fig. 4 M. speciosa (Murray, 1907). (a) Creeping, dorsal view (arrows pointing at skin fold); (b) and (c) Feeding, dorsal view (b. arrows pointing at cingulum and fold skin, c. arrows pointing at fold skin); (d) Feeding (head), dorsal view; (e) Creeping (foot), dorsal view; (f) Creeping (foot), lateral view; (g) Creeping (foot), ventral view |
表2 涠洲岛蛭态轮虫物种名录表Tab. 2 The species list of bdelloid rotifers in the Weizhou island |
| 科 | 属 | 种 | 生境类型 |
|---|---|---|---|
| 盘网轮科 Adinetidae | 盘网轮属Adineta | 急角盘网轮虫Adineta acuticornis Haigh, 1967 | 竹叶, 松树落叶, 其他落叶, 苔藓, 燃烧后的叶杆 |
| 贝式盘网轮虫A. beysunae Örstan, 2018 | 竹叶, 松树落叶 | ||
| 楔足盘网轮虫A. cuneata Milne, 1916 | 竹叶, 松树落叶, 其他落叶, 苔藓 | ||
| 游荡盘网轮虫A. vaga (Davis, 1873) | 竹叶, 松树落叶, 其他落叶, 苔藓, 燃烧后的叶杆 | ||
| 突盘轮Bradyscela | 跛足突盘轮虫Bradyscela clauda (Bryce, 1893) | 竹叶, 其他落叶 | |
| 宿轮科Habrotrochidae | 宿轮属 Habrotrocha | 双齿宿轮虫H. bidens (Gosse, 1851) | 竹叶, 松树落叶, 其他落叶, 苔藓 |
| 缢缩宿轮虫H. constricta (Dujardin, 1841) | 竹叶 | ||
| 领宿轮虫H. collaris Wulfert, 1965 | 竹叶, 苔藓 | ||
| 金色宿轮虫H. flava Bryce,1915 | 竹叶, 松树落叶 | ||
| 黄体宿轮虫H. flaviformis De Koning, 1947 | 竹叶, 其他落叶 | ||
| 舌凸宿轮虫H. ligula Bryce, 1913 | 松树落叶, 燃烧后的叶杆 | ||
| 罗莎宿轮虫H. rosa Donner, 1949 | 竹叶, 松树落叶, 其他落叶 | ||
| 三趾宿轮虫H. tripus (Murray, 1907) | 竹叶, 松树落叶, 其他落叶 | ||
| 弯唇宿轮虫H. thienemanni Hauer, 1924 | 竹叶 | ||
| 一种宿轮虫Habrotrocha sp.1 | 松树落叶 | ||
| 一种宿轮虫Habrotrocha sp.2 | 土壤 | ||
| 一种宿轮虫Habrotrocha sp.3 | 竹叶 | ||
| 一种宿轮虫Habrotrocha sp.4 | 竹叶 | ||
| 敖突轮属Otostephanos | 皇家敖突轮虫Otostephanos regalis Milne, 1916 | 其他落叶 | |
| 环颈敖突轮虫O. torquatus (Bryce, 1913) | 竹叶 | ||
| 旋轮科 Philodinidae | 粗颈轮属Macrotrachela | 短唇粗颈轮虫Macrotrachela brevilabris De Koning, 1947 | 松树落叶 |
| 精巧粗颈轮虫M. concinna (Bryce, 1912) | 竹叶, 苔藓 | ||
| 戈氏粗颈轮虫M. decora (Bryce, 1912)* | 竹叶, 土壤, 燃烧后的叶杆 | ||
| 欧氏粗颈轮虫M. ehrenbergii (Janson, 1893) | 竹叶, 松树落叶, 其他落叶, 苔藓 | ||
| 栖居粗颈轮虫M. habita (Bryce, 1894) | 竹叶, 松树落叶, 其他落叶, 苔藓, 土壤, 燃烧后的叶杆 | ||
| 毛粗颈轮虫M. induta Donner, 1951 | 其他落叶, 苔藓 | ||
| 无刺粗颈轮虫M. inermis Donner, 1965 | 竹叶, 松树落叶 | ||
| 奇异粗颈轮虫M. insolita De Koning, 1947 | 松树落叶 | ||
| 圆腹粗颈轮虫M. kallosoma (Schulte, 1954) | 竹叶 | ||
| 游离粗颈轮虫M. libera Donner, 1949 | 松树落叶, 土壤 | ||
| 多刺粗颈轮虫M. multispinosa (Murray, 1908) | 竹叶, 松树落叶, 其他落叶, 苔藓, 土壤, 带柄植物花瓣 | ||
| 舌凸矮小粗颈轮虫M. nana ligulata Schulte, 1954* | 竹叶 | ||
| 娜娜矮小粗颈轮虫M. nana nana (Bryce, 1912) | 竹叶, 松树落叶, 燃烧后的叶杆 | ||
| 四角粗颈轮虫M. quadricornifera Milne, 1886 | 竹叶, 松树落叶, 其他落叶, 苔藓, 带柄植物花瓣 | ||
| 美丽粗颈轮虫M. speciosa (Murray, 1907)* | 其他落叶 | ||
| 畏弱粗颈轮虫M. timida Milne, 1916 | 松树落叶 | ||
| 一种粗颈轮虫Macrotrachela sp.1 | 竹叶, 苔藓 | ||
| 一种粗颈轮虫Macrotrachela sp.2 | 竹叶 | ||
| 水蛭轮属Mniobia | 水蛭轮虫 Mniobia tentans Donner, 1949 | 竹叶, 松树落叶, 其他落叶, 苔藓 | |
| 一种水蛭轮虫Mniobia sp. | 竹叶 | ||
| 旋轮属Philodina | 宽沟锐角旋轮虫Philodina acuticornis odiosa Milne, 1916 | 苔藓, 带柄植物花瓣 | |
| 南极旋轮虫P. antarctica Murray, 1910 | 带柄植物花瓣 | ||
| 双戟旋轮虫P. duplicalcar (De Koning, 1947) | 其他落叶 | ||
| 巨环旋轮虫P. megalotrocha Ehrenberg, 1832 | 松树落叶 | ||
| 微戟旋轮虫P. parvicalcar De Koning, 1947 | 松树落叶, 苔藓, 苔藓土壤, 土壤 | ||
| 宽体旋轮虫P. plena (Bryce, 1894) | 竹叶, 松树落叶, 其他落叶, 苔藓, 土壤 | ||
| 迅捷旋轮虫P. rapida Milne, 1916 | 竹叶, 松树落叶, 其他落叶, 燃烧后的叶杆 | ||
| 褶皱旋轮虫P. rugosa Bryce, 1903 | 竹叶, 土壤 | ||
| 宁静旋轮虫P. tranquilla Wulfert, 1942 | 带柄植物花瓣 | ||
| 凶猛旋轮虫P. vorax (Janson, 1893) | 竹叶, 苔藓, 土壤 | ||
| 旋轮科 Philodinidae | 轮虫属Rotaria | 转轮虫R. rotatoria (Pallas, 1766) | 松树落叶 |
| 污轮虫R. sordida (Western, 1893) | 竹叶, 松树落叶, 其他落叶, 苔藓, 土壤 | ||
| 懒轮虫R. tardigrada (Ehrenberg, 1830) | 松树落叶 |
注: *为中国新记录种 |
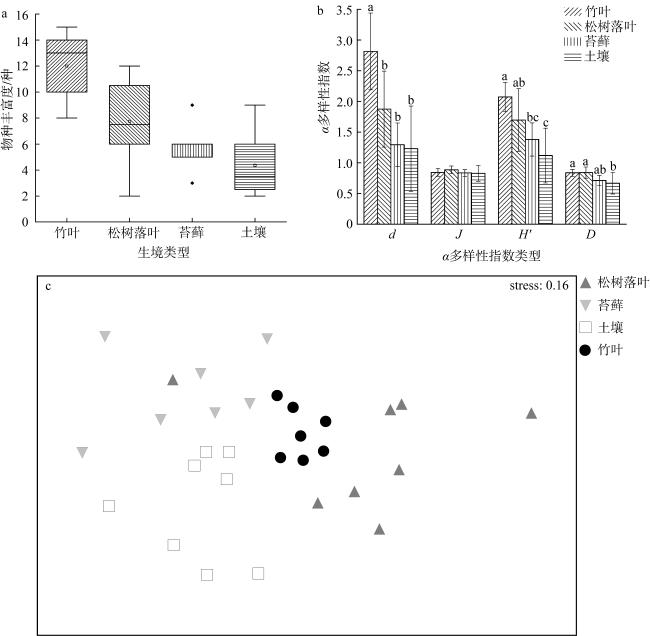
图5 涠洲岛4种生境物种丰富度箱型图(a), 涠洲岛4种生境α多样性指数图(b)和涠洲岛4种生境蛭态轮虫群落结构非度量多维尺度分析图(c)图b中d为Margalef丰富度指数, J为Pielou均匀度指数, H'为Shannon-Wiener指数, D为Simpson指数。图中不同小写字母表示生境间存在显著差异 Fig. 5 (a) Box plots of species richness of four habitats on the Weizhou island; (b) α diversity indices of four habitats on the Weizhou island; (c) Non-metric multidimensional scale analysis (NMDS) of bdelloid rotifer community structure in four habitats on the Weizhou island |
表3 4种生境物种丰富度差异的真实显著差异检验结果Tab. 3 Results of the Tukey HSD test for differences in species richness of the four habitats |
| 松树落叶 | 竹叶 | 苔藓 | |
|---|---|---|---|
| 竹叶 | 0.02461 | ||
| 苔藓 | 0.42002 | 0.00104 | |
| 土壤 | 0.08048 | <0.0001 | 0.86332 |
表4 蛭态轮虫4种生境丰度、IndVal指数、p值和生境偏好Tab.4 Abundance, IndVal value, p value and habitat preference of bdelloid rotifers in four habitats |
| 物种 | 丰度 | IndVal/% | p 值 | 生境偏好 | |||
|---|---|---|---|---|---|---|---|
| 竹叶 | 松树落叶 | 苔藓 | 土壤 | ||||
| 游荡盘网轮虫(A. vaga) | 8 | 45 | 1 | 0 | 72.3 | 0.013 | 竹叶, 松树落叶 |
| 双齿宿轮虫(H. biddens) | 15 | 16 | 0 | 0 | 68.3 | 0.022 | 竹叶, 松树落叶 |
| 金色宿轮虫(H. flava) | 13 | 6 | 0 | 0 | 63.8 | 0.043 | 竹叶 |
| 罗莎宿轮虫(H. rosa) | 13 | 51 | 0 | 0 | 89.4 | 0.001 | 竹叶, 松树落叶 |
| 三趾宿轮虫(H. tripus) | 9 | 19 | 0 | 0 | 68.3 | 0.022 | 竹叶, 松树落叶 |
| 戈氏粗颈轮虫(M. decora) | 1 | 0 | 3 | 32 | 81.0 | 0.006 | 土壤 |
| 多刺粗颈轮虫(M. multispinosa) | 94 | 8 | 28 | 56 | 80.9 | 0.027 | 竹叶, 苔藓, 土壤 |
| 四角粗颈轮虫(M. quadricornifera) | 29 | 1 | 26 | 1 | 74.4 | 0.017 | 竹叶, 苔藓 |
| 宽体旋轮虫(P. plena) | 31 | 16 | 0 | 3 | 79.4 | 0.004 | 竹叶, 松树落叶 |
| 迅捷旋轮虫(P. rapida) | 8 | 22 | 0 | 0 | 68.3 | 0.023 | 竹叶, 松树落叶 |
表5 蛭态轮虫4种生境优势种及优势度Tab. 5 The dominant species and dominance of bdelloid rotifers in four habitats |
| 种类组成 | 区域优势度 | 竹叶 | 松树落叶 | 苔藓 | 土壤 |
|---|---|---|---|---|---|
| 急角盘网轮虫(A. acuticornis) | / | 0.03 | / | 0.03 | / |
| 游荡盘网轮虫(A. vaga) | / | / | 0.06 | / | — |
| 双齿宿轮虫(H. bidens) | / | 0.03 | / | — | — |
| 金色宿轮虫(H. flava) | / | 0.02 | / | — | — |
| 罗莎宿轮虫(H. rosa) | 0.02 | 0.03 | 0.13 | — | — |
| 三趾宿轮虫(H. tripus) | / | / | 0.02 | — | — |
| 戈氏粗颈轮虫(M. decora) | / | / | — | / | 0.11 |
| 栖居粗颈轮虫(M. habita) | 0.03 | / | / | 0.19 | 0.03 |
| 多刺粗颈轮虫(M. multispinosa) | 0.09 | 0.27 | / | 0.07 | 0.16 |
| 四角粗颈轮虫(M. quadricornifera) | / | 0.07 | / | 0.03 | / |
| 一种水蛭轮虫(Mniobia sp.) | / | — | / | 0.05 | — |
| 微戟旋轮虫(P. parvicalcar) | / | — | / | 0.02 | / |
| 宽体旋轮虫(P. plena) | / | 0.08 | / | — | / |
| 迅捷旋轮虫(P. rapida) | / | / | 0.02 | — | — |
| 凶猛旋轮虫(P. vorax) | / | / | — | — | 0.03 |
| 污轮虫(R. sordida) | 0.07 | 0.09 | / | 0.03 | 0.27 |
注: “/”表示优势度< 0.02, “—”表示未出现 |
| [1] |
王庆, 李莹, 汪文博, 等, 2021. 广东省蛭态轮虫物种多样性及其系统发育研究[J]. 中国环境科学, 41(9): 4367-4377.
|
| [2] |
汪文博, 李莹, 王晓妍, 等, 2022. 中国蛭态轮虫亚纲6种新记录种记述[J]. 动物学杂志, 57(4): 595-606.
|
| [3] |
汪文博, 王庆, 李莹, 等, 2021. 落叶生境蛭态轮虫物种多样性及四种中国新记录种[J]. 水生生物学报, 45(2): 436-445.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}