
西沙永乐环礁造礁石珊瑚共生体对低光环境的生理响应
|
王永智(1996—), 男, 吉林省通化市人, 工程师, 从事珊瑚礁生态学研究。email: |
Copy editor: 林强
收稿日期: 2023-07-03
修回日期: 2023-08-21
网络出版日期: 2023-09-04
基金资助
国家自然科学基金项目(42277362)
国家自然科学基金项目(41806139)
广州市科技计划项目(202002030345)
中央级公益性科研院所基本科研业务专项(PM-zx703-202105-176)
中央级公益性科研院所基本科研业务专项(PM-zx703-202004-143)
Physiological responses to light limitation of reef-building corals in the Yongle Atoll of the Xisha Islands
Copy editor: LIN Qiang
Received date: 2023-07-03
Revised date: 2023-08-21
Online published: 2023-09-04
Supported by
National Natural Science Foundation of China(42277362)
National Natural Science Foundation of China(41806139)
Science and Technology Planning Project of Guangzhou City(202002030345)
Central Public-Interest Scientific Institution Basal Research Fund(PM-zx703-202105-176)
Central Public-Interest Scientific Institution Basal Research Fund(PM-zx703-202004-143)
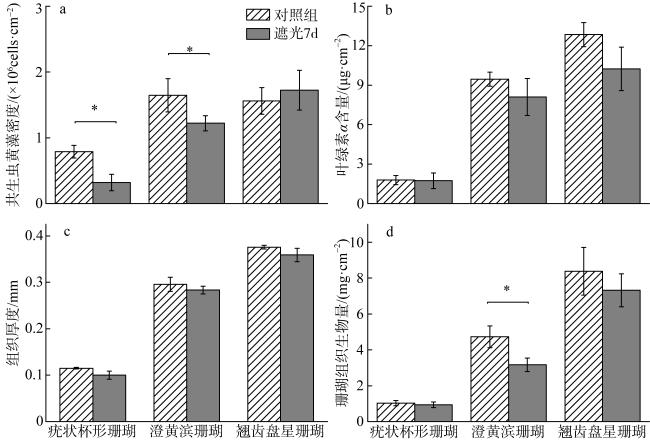
光照是影响造礁石珊瑚生长与分布的重要因素之一, 但目前关于造礁石珊瑚对低光环境的响应特征与适应性尚不清楚。本研究以西沙常见的疣状杯形珊瑚(Pocillopora verrucosa)、澄黄滨珊瑚(Porites lutea)和翘齿盘星珊瑚(Dipsastraea matthaii)为研究对象, 于2021年春季在永乐环礁10m水深处开展了为期1周的原位遮光实验, 其中遮光实验组与正常对照组相比光强削减了80%~90%, 其日间有效光辐射(photosynthetically active radiation, PAR)为20.2~41.5μmol photons·m-2·s-1。实验结果显示, 弱光状态下疣状杯形珊瑚、澄黄滨珊瑚和翘齿盘星珊瑚的实际光量子产量(ΦPSII)明显升高, 最大电子传递速率(r-ETRmax)和饱和辐射光强(Ek)出现了下降的现象, 同时三种珊瑚的共生藻密度(翘齿盘星珊瑚除外)、叶绿素a含量、组织生物量和组织厚度出现了不同程度的下降, 其中以澄黄滨珊瑚的变化最为显著, 其共生藻密度下降了14.2%, 组织生物量下降了32.9%。短期弱光限制中,造礁石珊瑚能够通过提高自身的实际光合效率或(和)降低虫黄藻密度适应光强的不足, 但珊瑚共生体通过光合作用转化的能量短期内无法满足珊瑚宿主自身正常生长的代谢需求, 因而出现消耗大于获得、珊瑚组织生物量和组织厚度短暂下降的情况。
王永智 , 许莉佳 , 黄柏强 , 杨天件 , 綦世斌 , 陈辉 , 杨静 . 西沙永乐环礁造礁石珊瑚共生体对低光环境的生理响应[J]. 热带海洋学报, 2024 , 43(3) : 31 -39 . DOI: 10.11978/2023092
Light is one of the most important factors influencing the growth and distribution of reef corals, but the physiological response and adaptability of reef corals to low light is not well understood. In this study, we used an in-situ shading experiment with three typical coral species (Pocillopora verrucosa, Porites lutea and Dipsastraea matthaii) to test how physiological response of corals under light limitation. The experiment site is located at the depth of 10 meters of the Yongle atoll, in which the irradiance in the shaded area was reduced by 80%~90%, and the photosynthetically active radiation (PAR) ranges from 20.2 μmol photons·m-2·s-1 to 41.5 μmol photons·m-2·s-1. The results showed that the physiological parameters of three corals changed significantly under light limitation. During 7-day experiments, the effective photosynthetic efficiency (Φ PSII) of the three corals increased, while the maximum electron transport rate (r-ETRmax) and the minimum saturating irradiance (Ek) decreased significantly. Meanwhile, the zooxanthellae density (except Dipsastraea matthaii), chlorophyll a content, tissue thickness and tissue biomass of the three corals decreased in diverse degrees, with the most severe reduction found in Porites lutea that the zooxanthellae density and tissue biomass decreased by 14.2%and 32.9%, respectively. These results suggest that the reef corals adapt to shading by improving their effective photosynthetic efficiency in short-term light limitation, but the energy transformed by photosynthesis could not meet the metabolic needs of coral growth, thus coral tissue biomass and tissue thickness decrease.
Key words: scleractinia coral; light limitation; photo-physiology; Yongle Atoll
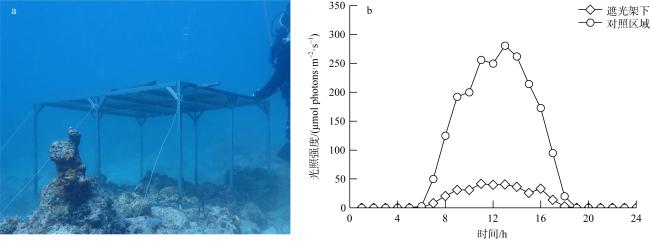
图1 珊瑚遮光原位实验照片(a)和遮光区及对照区光照强度日变化(b)Fig. 1 Graph of the in-situ light-shading experiment in the Yongle Atoll (a), and the diurnal changes of photosynthetically active radiation in areas with or without shading (b) |
表1 实验珊瑚礁区海水环境因子Tab. 1 Seawater environmental parameters in the experiment coral reefs |
| 环境因子 | 平均值±标准差 |
|---|---|
| SST/℃ | 25.7±0.2 |
| 盐度/‰ | 33.68±0.28 |
| pH | 8.11±0.01 |
| 溶解氧含量/(mg·L-1) | 6.56±0.17 |
| 叶绿素 a含量/(μg·L-1) | 0.18±0.04 |
| 无机氮含量/(μmol·L-1) | 1.46±0.31 |
| 无机磷含量/(μmol·L-1) | 0.05±0.01 |
表2 荧光参数意义与计算方法Tab. 2 Parameters and definitions of fluorescence |
| 荧光参数 | 计算公式 | 意义 | |
|---|---|---|---|
| F0 | 初始荧光, 即在暗适应状态下当 PSII 的所有反应中心处于完全开放状态并且所有的非光化学过程处于最小时的荧光产量 | ||
| F0’ | 光适应状态下的初始荧光 | ||
| Fm | 最大荧光, 即在暗适应状态下当 PSII 的所有反应中心处于完全关闭状态并且所有的非光化学过程处于最大时的荧光产量 | ||
| Fm’ | 光适应状态下的最大荧光 | ||
| Ft | 任意时间实际荧光产量 | ||
| ΦPSII | (Fm’-Ft)/Fm’ | PSII 的实际光量子产量, 反映造礁石珊瑚实时的光合速率 | |
| qP | (Fm’-Ft) /(Fm’-F0’) | 光化学淬灭系数, 即激发能被开放的反应中心捕获并转化为化学能而导致的荧光淬灭, 反映了光适应状态下造礁石珊瑚的光能利用转化能力 | |
| NPQ | (Fm - Fm’)/ Fm’ | 非光化学淬灭系数, 即激发能以热能形式耗散而导致的荧光淬灭, 反映光适应状态下造礁石珊瑚在光能利用中热耗散能量的比例 | |
| Y(NPQ) | Ft/ Fm’- Ft/ Fm | 调节性能量耗散量子产量, 反映造礁石珊瑚在光适应状态下光合过程中非利用光能自我调节性耗散的比例 | |
| Y(NO) | F/ Fm | 非调节性能量耗散量子产量, 反映造礁石珊瑚在光适应状态下的光损伤情况 | |
| r-ETRmax | 通过LC曲线拟合获取 | 最大电子传递速率, 反映造礁石珊瑚在饱和光强下的光能利用能力 | |
| Ek | r-ETRmax/LC曲线初始斜率 | 饱和辐射光强, 反映造礁石珊瑚能够有效利用的最大光照强度 |
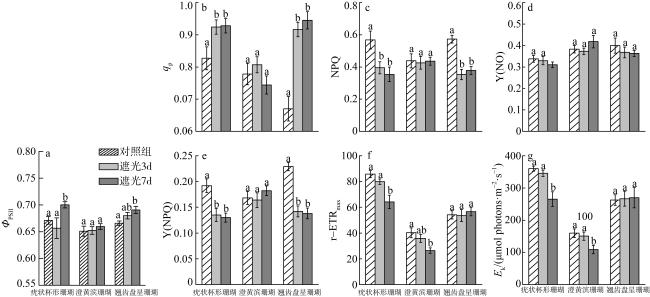
图2 遮光实验中三种珊瑚的荧光参数a. 实际光量子产量ΦPSII; b. 光化学淬灭系数qP; c. 非光化学淬灭系数NPQ; d. 调节性能量耗散量子产量Y(NO); e. 非调节性能量耗散的量子产量Y(NPQ); f. 最大相对电子传递速率r-ETRmax; g. 饱和光强Ek。柱状图内不同字母标记表明存在显著性差异(P<0.05) Fig. 2 The effect of shading on fluorescence parameters of coral. (a)ΦPSII; (b) qP; (c) NPQ; (d) Y(NO); (e)Y(NPQ); (f) r-ETRmax; (g) Ek. letters above histograms denote statistical differences (P<0.05) |
| [1] |
何茜, 俞晓磊, 梁宇娴, 等, 2022. 升温对丛生盔形珊瑚两种形态型代谢和共生藻光合生理的影响[J]. 热带海洋学报, 41(5): 133-140.
|
| [2] |
黄晖, 俞晓磊, 雷新明, 等, 2020. 环境变化对造礁石珊瑚营养方式的影响及其适应性[J]. 海洋科学进展, 38(2): 189-198.
|
| [3] |
江绿苗, 陈天然, 赵宽, 等, 2023. 南海北部涠洲岛边缘珊瑚礁的生物侵蚀实验研究[J/OL]. 热带海洋学报. [2023-04-17]. https://doi.org/10.11978/2023002.
|
| [4] |
门征, 陈汉吉, 许慎栋, 等, 2023. 珊瑚脂质是其共生虫黄藻密度降低时的重要能量来源[J]. 海洋学报, 45(1): 71-79.
|
| [5] |
赵美霞, 余克服, 张乔民, 等, 2009. 近50a来三亚鹿回头石珊瑚物种多样性的演变特征及其环境意义[J]. 海洋环境科学, 28(2): 25-130.
|
| [6] |
中国国家标准化管理委员会, 2007. 海洋调查规范第4部分:海水化学要素调查, GB/T 12763. 4-2007[S]. 北京: 中国标准出版社: 1-65.
Standardization Administration of the People's Republic of China, 2007, Specifications for oceanographic survey part: Survey of chemical parameters in sea water, GB/T 12763. 4-2007[S]. Beijing: Standards Press of China: 1-65 (in Chinese).
|
| [7] |
周洁, 施祺, 余克服, 2011. 叶绿素荧光技术在珊瑚礁研究中的应用[J]. 热带地理, 31(2): 223-229.
|
| [8] |
周洁, 余克服, 施祺, 2014. pCO2增加引起的海洋酸化对造礁珊瑚光合效率的影响[J]. 海洋与湖沼, 45(1): 39-51.
|
| [9] |
朱文涛, 夏景全, 刘相波, 等, 2022. 丛生盔形珊瑚光合生理及共生真菌群落分析[J]. 热带海洋学报, 41(2): 132-141.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}