
|
黄晖(1969—), 女, 江西省分宜县人, 研究员, 从事珊瑚生物学与珊瑚礁生态学等方面研究。email: |
Copy editor: 林强
收稿日期: 2023-07-25
修回日期: 2023-09-01
网络出版日期: 2023-09-06
基金资助
国家自然科学基金项目(41976120)
国家自然科学基金项目(42276124)
国家重点研发计划项目(2021YFC31005001)
Current status and prospects of coral reef ecology research
Copy editor: LIN Qiang
Received date: 2023-07-25
Revised date: 2023-09-01
Online published: 2023-09-06
Supported by
National Natural Science Foundation of China(41976120)
National Natural Science Foundation of China(42276124)
National Key Research and Development Program(2021YFC31005001)
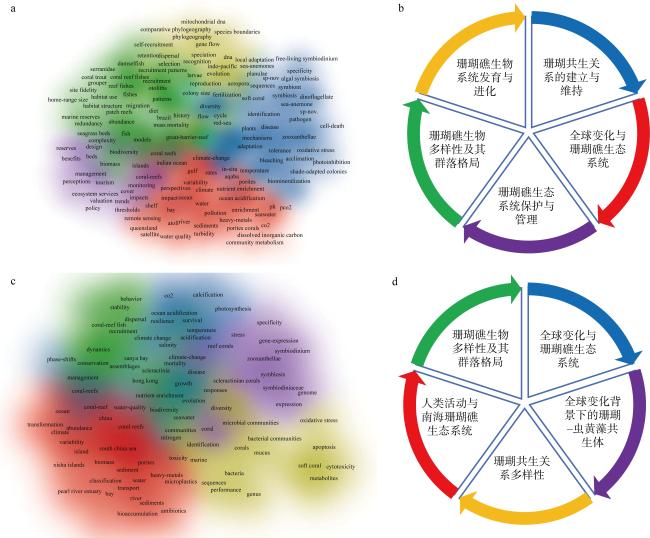
珊瑚礁生态系统是海洋生态系统的重要组成部分, 在维持海洋生态平衡和生物多样性上扮演着重要角色。本文通过文献检索手段, 归纳了当前国际和国内珊瑚礁生态学领域的发展现状。国际上关注的主要方面包括全球变化与珊瑚礁生态系统、珊瑚共生关系的建立与维持、珊瑚礁生物系统发育与进化、珊瑚礁生物多样性及其物种分布格局、珊瑚礁生态系统保护与管理等; 国内关注的主要方面包括全球变化与珊瑚礁生态系统、全球变化背景下的珊瑚-虫黄藻共生体、珊瑚共生关系多样性、人类活动与南海珊瑚礁生态系统、珊瑚礁生物多样性及其群落格局等。珊瑚礁生态学研究的未来发展趋势将更加注重全球变化背景下的珊瑚与珊瑚礁生态系统、珊瑚共生机制、边缘珊瑚礁以及珊瑚礁保护与修复。未来我国珊瑚礁生态学研究需提升全球视野, 加强国内与国际合作, 深入机制挖掘, 完善系统研究, 增强珊瑚礁保护与修复研究。
黄晖 , 俞晓磊 , 黄林韬 , 江雷 . 珊瑚礁生态学研究现状和展望[J]. 热带海洋学报, 2024 , 43(3) : 3 -12 . DOI: 10.11978/2023101
Coral reef ecosystems are important components of marine ecosystems and play a crucial role in maintaining marine ecological balance and biodiversity. This article summarizes the current international and domestic development status of coral reef ecology through literature review. The major international research focuses include global change and coral reef ecosystems, establishment and maintenance of coral symbiosis, development and evolution of coral reef biotic systems, coral reef biodiversity and species distribution patterns, as well as coral reef ecosystem conservation and management. The major domestic research focuses include global change and coral reef ecosystems, coral-algal symbiosis under global change, diversity of coral symbiotic relationships, human activities and the South China Sea coral reef ecosystem, and coral reef biodiversity and community patterns. The future development trends in coral reef ecology research will emphasize the coral and coral reef ecosystems under the context of global change, coral symbiotic mechanisms, marginal coral reefs, and coral reef conservation and restoration. Future research in coral reef ecology in China should enhance global perspectives, strengthen domestic and international collaborations, explore underlying mechanisms, improve systematic research, and enhance coral reef conservation and restoration studies.
图1 珊瑚礁生态学研究动态a. 国际珊瑚礁生态学研究高频关键词; b. 国际珊瑚礁生态学研究热点; c. 国内珊瑚礁生态学研究高频关键词; d. 国内珊瑚礁生态学研究热点 Fig. 1 Research trends in coral reef ecology. (a) High-frequency keywords for international coral reef ecology research; (b) international research hotspots in coral reef ecology; (c) high-frequency keywords for domestic coral reef ecology research; (d) domestic research hotspots in coral reef ecology |
| [1] |
黄晖, 陈竹, 黄林韬, 2021. 中国珊瑚礁状况报告(2010-2019)[M]. 北京: 海洋出版社.
|
| [2] |
施祺, 赵美霞, 黄玲英, 等, 2010. 三亚鹿回头岸礁区人类活动及其对珊瑚礁的影响[J]. 热带地理, 30(5): 486-490, 509.
|
| [3] |
王耕, 常畅, 于小茜, 等, 2019. 基于文献计量分析的珊瑚礁研究现状与热点[J]. 生态学报, 39(3): 1114-1123.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}