
南沙群岛珊瑚礁区黑斑鹦嘴鱼(Scarus globiceps)食性分析
Copy editor: 林强
网络出版日期: 2023-09-05
Diet analysis of the parrotfish (Scarus globiceps) in coral reefs of the Nansha Islands
Copy editor: LIN Qiang
Online published: 2023-09-05
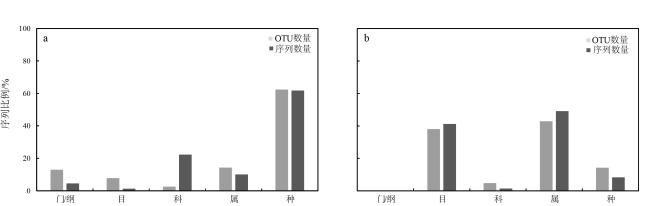
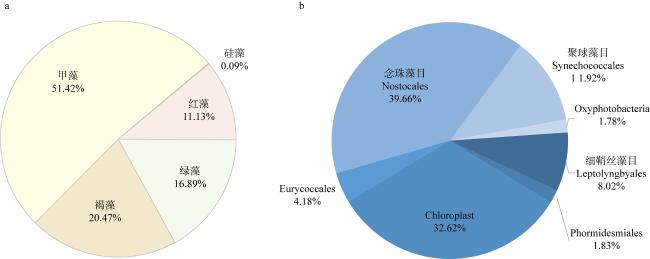
鹦嘴鱼科(Scaridae)鱼类参与珊瑚礁生态系统诸多关键生态过程, 在维持珊瑚礁生态系统稳定与平衡中发挥着重要作用。由于以往研究手段的限制, 对鹦嘴鱼的食物来源及生态功能价值认识不足, 在其功能定位方面存在较多争议。本研究选择珊瑚分布的典型区域—南沙群岛中的东门礁和南薰礁为研究海域, 对该海域鹦嘴鱼优势种类黑斑鹦嘴鱼(Scarus globiceps)摄食的藻类多样性进行全面分析。通过18S rDNA和16S rDNA 多基因条形码技术, 分别对黑斑鹦嘴鱼肠含物的真核藻类和原核藻类DNA进行高通量测序分析。18S rDNA的测序结果发现, 黑斑鹦嘴鱼类的肠含物中真核藻类以甲藻(Dinoflagellata)、红藻(Rhodophyta)、绿藻(Chlorophyta)、褐藻(Ochrophyta)为主, 共计77个OTU(operational taxonomic units)。甲藻相对序列丰度和多样性较高, 序列比例占真核藻类序列总数的51.42%, 其中网甲藻科(Suessiaceae)OTU_5在东门礁和南薰礁的样品中均超过20%。16S rDNA测序鉴定发现肠含物含有原核藻类(蓝藻)的序列, 共计21个OTU, 其中以念珠藻目(Nostocales)的相对序列丰度最高, 达到39.33%。本研究表明, 黑斑鹦嘴鱼在摄食过程中会摄入一定量大型藻类, 但微藻(甲藻)序列占据优势地位, 蓝藻在肠含物中也有较高的检出率, 说明需要重新考虑微藻(甲藻和蓝藻)在鹦嘴鱼食源中的重要贡献以及对珊瑚礁生态系统结构与功能的影响。
林先智 , 周岩岩 , 林皓晔 , 胡思敏 , 黄晖 , 张黎 , 刘胜 . 南沙群岛珊瑚礁区黑斑鹦嘴鱼(Scarus globiceps)食性分析[J]. 热带海洋学报, 2024 , 43(3) : 100 -108 . DOI: 10.11978/2023102
The parrotfish (Scaridae) plays an important role in maintaining the stability and balance of coral reef ecosystems, participating in many key ecological processes. Due to limitations in previous research methods, there is insufficient understanding of the food sources and ecological functional value of parrotfish, leading to controversies regarding their functional role. In this study, the typical areas of coral distribution, the Dongmen Reef and the Nanxun Reef in the Nansha Islands, were selected as the study sites, and the algal diversity consumed by the dominant species of parrotfish, Scarus globiceps, in this region was comprehensively analyzed. High-throughput sequencing analysis of the DNA in the intestinal contents of S. globiceps was conducted using both 18S rDNA and 16S rDNA. The sequencing results of 18S rDNA revealed that the intestinal contents of S. globiceps consisted mainly of several types of eukaryotic algae, including Dinoflagellata, Rhodophyta, Chlorophyta, and Ochrophyta, with a total of 77 operational taxonomic units (OTUs). Dinoflagellata had relatively high sequence abundance and diversity, accounting for 51.42% of the total eukaryotic algae sequences, with one species from the family Suessiaceae exceeding 20% in both the Dongmen Reef and Nanxun Reef samples. The sequencing of 16S rDNA identified sequences of prokaryotic algae (cyanobacteria) in the intestinal contents, with a total of 21 OTUs, and the relative sequence abundance of the order Nostocales was the highest, reaching 39.33%. This study indicates that although S. globiceps consumes a certain amount of macroalgae during feeding, microalgae (dinoflagellates and cyanobacteria) still dominate, suggesting the need to reconsider the important contribution of microalgae to the diet of parrotfish and their impact on the structure and function of coral reef ecosystems.
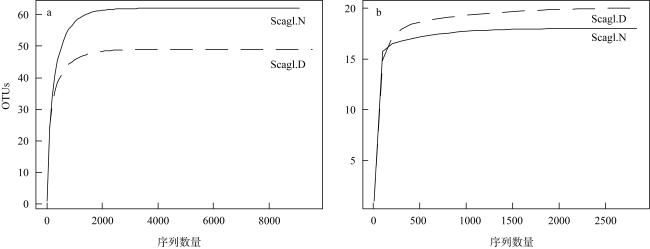
表1 黑斑鹦嘴鱼采样信息Tab. 1 The sampling information of parrotfish |
| 编号 | 站点 | 取样数 | 体重/g | 体长/cm |
|---|---|---|---|---|
| Scagl. D | 东门礁 | 8 | 60.33±17.04 | 11.91±1.15 |
| Scagl. N | 南薰礁 | 8 | 55.03±26.33 | 11.61±2.11 |
| [1] |
林先智, 胡思敏, 刘胜, 等, 2018. 传统测序与高通量测序在稚鱼食性分析中的比较[J]. 应用生态学报, 29(9): 3093-3101.
|
| [2] |
张琛, 胡思敏, 林先智, 等, 2022. 南沙珊瑚礁区波纹钩鳞鲀(Balistapus undulatus)食性和营养级分析[J]. 热带海洋学报, 41(1): 7-14.
|
| [3] |
周天成, 胡思敏, 林先智, 等, 2020. 基于18S rDNA条形码技术的珊瑚礁区塔形马蹄螺(Tectus pyramis)食性分析[J]. 海洋科学, 44(2): 99-107.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}