
红树林生态重建效果预测研究新模式
|
吴鸿博(2000—), 男, 江西南昌人, 硕士研究生, 主要从事河口海岸数值模拟研究。email: |
Copy editor: 孙翠慈
收稿日期: 2023-06-26
修回日期: 2023-09-11
网络出版日期: 2023-10-09
基金资助
国家重点研发计划项目(2022YFC3106100)
江苏省海洋科技创新项目(JSZRHYKJ202105)
南通社会民生科技计划项目(MS12022009)
南通社会民生科技计划项目(MS22022082)
南通社会民生科技计划项目(MS22022083)
A novel pattern for predicting the effects of mangrove ecological reconstruction
Copy editor: SUN Cuici
Received date: 2023-06-26
Revised date: 2023-09-11
Online published: 2023-10-09
Supported by
National Key Research and Development Program of China(2022YFC3106100)
Marine Science and Technology Innovation Project of Jiangsu Province(JSZRHYKJ202105)
Nantong Science and Technology Bureau(MS12022009)
Nantong Science and Technology Bureau(MS22022082)
Nantong Science and Technology Bureau(MS22022083)
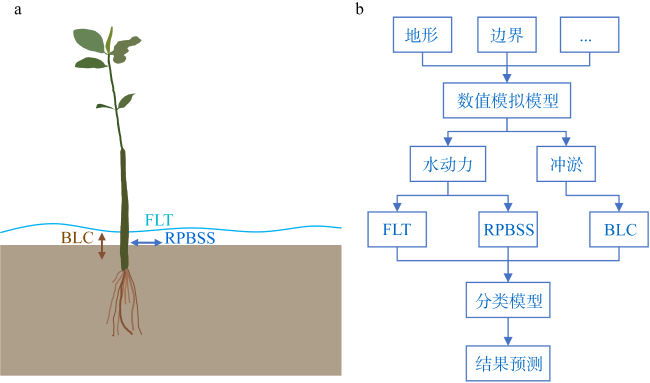
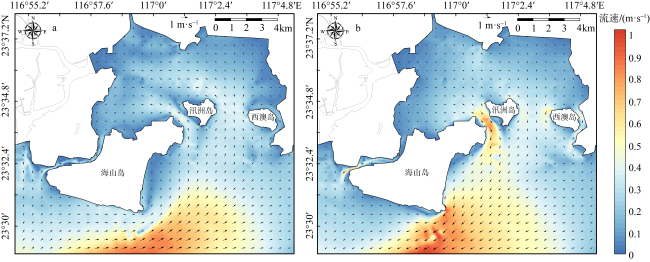
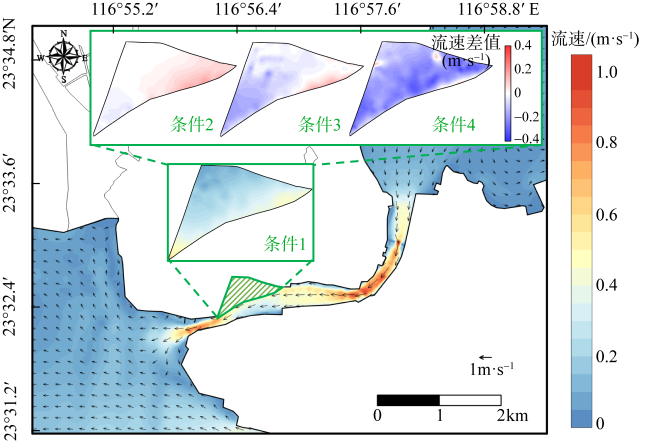
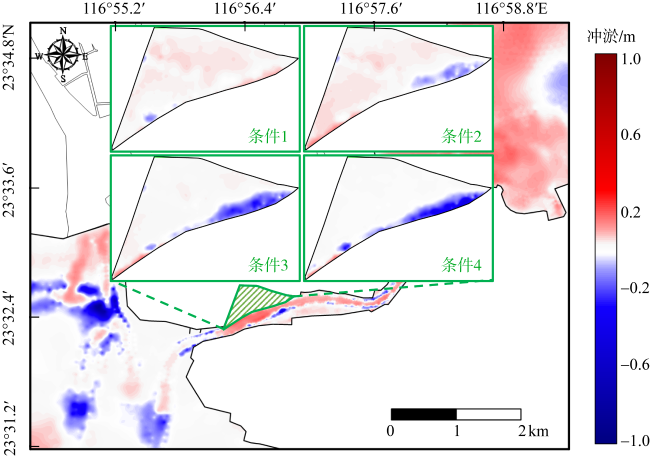
红树林是生态文明建设的组成部分, 对实现碳中和目标意义重大, 近年来实施了大量的保护和修复工程。然而, 目前缺少一种预测方法来比选有效的保护措施, 以指导红树林的生态重建工作。因此, 提出了一种“数值模拟模型+分类模型”的研究新模式。在模式应用中, 模拟了广东省柘林湾一年的水沙动力环境。通过考虑淹没时间、相对峰值底床切应力和底床高程变化, 描述了红树定植过程中所受到的胁迫, 对4种条件下笠港潮汐汊道内侧滩涂红树林的生态重建效果做出了预测。结果表明, 恢复汊道水深会轻微改变区域的水动力和泥沙冲淤环境, 导致生态重建成功率降低了3.12%; 提高区域底床高程会显著减小流速并加剧外侧冲刷, 提高0.5m和1.0m的高程分别使生态重建成功率提升了16.43%和32.75%。
吴鸿博 , 罗锋 , 陈治澎 , 朱飞 , 曾靖伟 , 张弛 , 李瑞杰 . 红树林生态重建效果预测研究新模式[J]. 热带海洋学报, 2024 , 43(4) : 86 -97 . DOI: 10.11978/2023088
Mangroves are an integral part of ecological civilization and hold significant importance in achieving carbon neutrality goals. In recent years, numerous conservation and restoration efforts have been undertaken to protect mangrove ecosystems. However, there is currently a lack of predictive methods for comparing and selecting effective conservation measures to guide mangrove ecological reconstruction. Therefore, a novel research approach combining “numerical simulation models” with “classification models” is proposed. In the application of this approach, the hydrodynamic and sedimentary environment of the Zhelin Bay in Guangdong Province was simulated for one year. By considering factors such as flooding time, relative peak bed shear stress, and bed level change, the stressors faced during the process of mangrove colonization were described. And ecological reconstruction effects of mangroves distributed on the inner tidal flats of the Ligang tidal inlet were predicted under four different conditions. The results indicate that restoring the inlet depth slightly alters the regional hydrodynamics and sediment environment, leading to a decrease of 3.12% in the success rate of ecological reconstruction. Elevating the bed elevation significantly reduces flow velocity and intensifies outer-side scouring. Raising the elevation by 0.5 meters and 1.0 meter increases the success rate of ecological reconstruction by 16.43% and 32.75%, respectively.
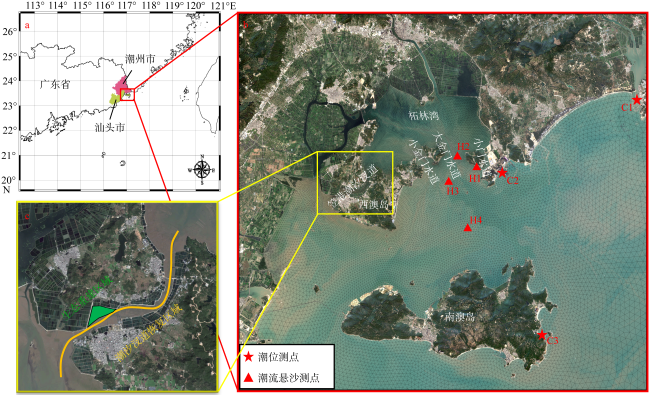
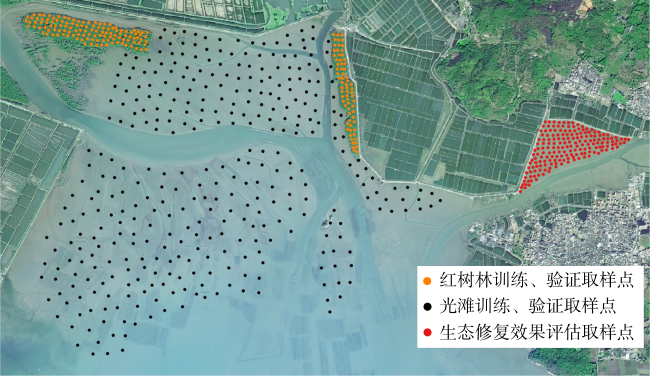
图2 研究区域位置图图2a 基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2023)2765号的标准地图制作; a. 研究区域的总体方位; b. 柘林湾及周边水道、岛屿位置, 各测点位置; c. 生态重建区域及潮汐汊道恢复区域位置 Fig. 2 Location of the study area. (a) General location of the study area; (b) location of the Zhelin Bay and surrounding waterways and islands, location of each measuring points; (c) location of the ecological reconstruction area and tidal inlet restoration area |
表1 模型输入的高程条件Tab. 1 Elevation conditions for model inputs |
| 高程条件设置 | 潮汐汊道恢复区域底床高程 | 生态修复区域滩面高程 |
|---|---|---|
| 条件1 | 原始高程 | 原始高程 |
| 条件2 | -3.0m | 原始高程 |
| 条件3 | -3.0m | 加高0.5m |
| 条件4 | -3.0m | 加高1.0m |
表2 模型部分参数Tab. 2 Part of model parameters |
| 参数 | 取值 | 单位 |
|---|---|---|
| 曼宁系数 | 0.0167 | m-1/3·s |
| 沉降速度系数 | 10 | m·s-1 |
| 临界沉积切应力 | 0.12 | N·m-2 |
| 侵蚀系数 | 5×10-5 | kg·s·m-2 |
| 临界侵蚀切应力 | 0.25 | N·m-2 |
| 底床密度 | 800 | kg ·m-3 |
| 底床糙率 | 0.0001 | m |
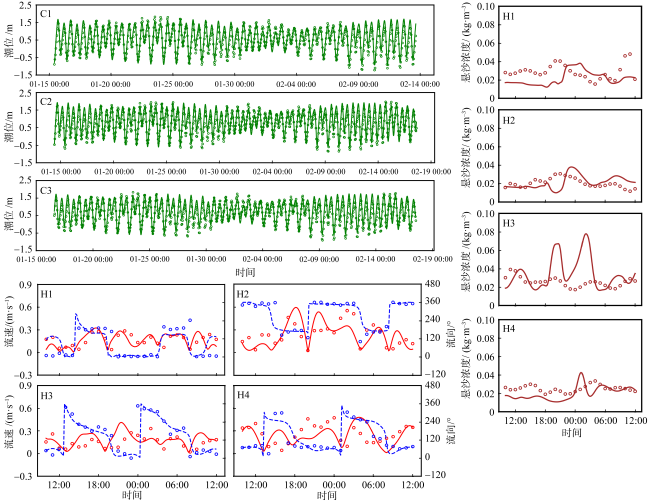
图3 水沙动力模型验证图(潮位、流速与流向、悬沙浓度)图中点代表实测数据, 而线代表模型模拟数据的拟合曲线。其中, 在流速流向图中, 实线代表流速, 虚线代表流向 Fig. 3 Hydrodynamic and sediment dynamic model validation (tide level; flow velocity and flow direction; suspended sediment concentration). The points in the figures represent measured data, while the lines represent the fitted curves of the model simulation data. In the figures of flow velocity and flow direction, solid lines represent flow velocity, and dashed lines represent flow direction |
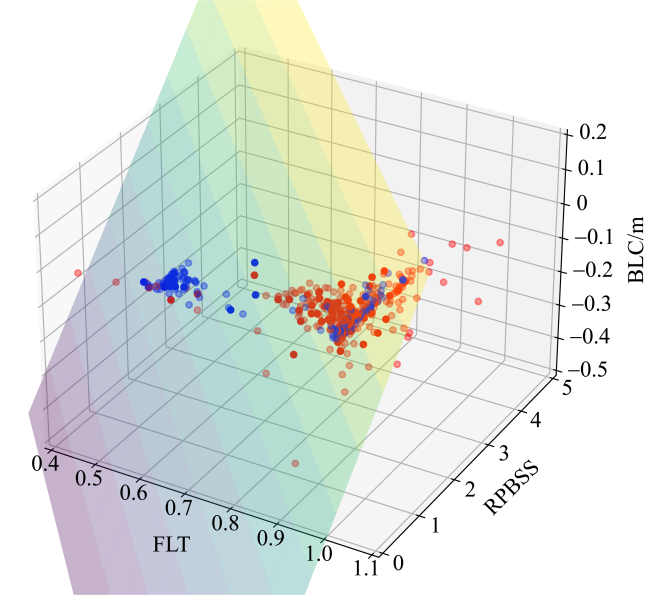
图8 逻辑回归训练集和验证集三维分布图蓝色点表示红树林取样点, 红色点表示光滩取样点, 不透明点表示分类正确, 半透明点表示分类错误 Fig. 8 The 3D distribution of the training and validation sets of logistic regression. Blue dots represent mangrove sampling points, and red dots represent mudflat sampling points. Solid dots indicate correct classification, while translucent dots indicate misclassification |
| [1] |
蔡爱智, 1994. 粤东柘林湾的泥沙来源与沉积环境[J]. 厦门大学学报(自然科学版), 33(4): 515-520.
|
| [2] |
陈鹭真, 王文卿, 林鹏, 2005. 潮汐淹水时间对秋茄幼苗生长的影响[J]. 海洋学报, 27(2): 141-147.
|
| [3] |
陈鹭真, 林鹏, 王文卿, 2006. 红树植物淹水胁迫响应研究进展[J]. 生态学报, 26(2): 586-593.
|
| [4] |
郭旭东, 2017. 全球变化下外来红树植物无瓣海桑和本土红树植物秋茄的响应情况[D]. 厦门: 厦门大学.
|
| [5] |
黄凡非, 唐丽丽, 李瑞利, 2023. 近四十年来中国大陆红树林保护与恢复措施的生态效益分析[J]. 北京大学学报(自然科学版), 59(5): 813-822.
|
| [6] |
蒋睢耀, 1993. 三百门港泥沙淤积研究[J]. 水道港口, (3): 28-35.
|
| [7] |
欧素英, 罗凯文, 田枫, 2016. 柘林湾多口门潮汐汊道动力地貌的演变[J]. 热带海洋学报, 35(2): 83-92.
|
| [8] |
潘炜杰, 祝振昌, 蔡宴朋, 等, 2021. 潮滩冲淤扰动下外来与乡土红树植物幼苗稳定性差异[J]. 热带海洋学报, 40(6): 120-127.
|
| [9] |
杨克用, 1990. 汕头柘林湾港区航道浚深后的回淤分析与计算[J]. 海岸工程, 9(3-4): 72-80.
|
| [10] |
张乔民, 赵焕庭, 宋朝景, 1991. 华南海岸的汇潮水道[M]// 中国地理学会地貌与第四纪专业委员会. 地貌及第四纪研究进展. 北京: 测绘出版社: 211-217 (in Chinese).
|
| [11] |
张乔民, 于红兵, 陈欣树, 等, 1997. 红树林生长带与潮汐水位关系的研究[J]. 生态学报, 17(3): 258-265.
|
| [12] |
张乔民, 隋淑珍, 张叶春, 等, 2001. 红树林宜林海洋环境指标研究[J]. 生态学报, 21(9): 1427-1437.
|
| [13] |
自然资源部,国家林业和草原局, 2020. 自然资源部国家林业和草原局关于印发《红树林保护修复专项行动计划(2020-2025年)》的通知[R]. 北京: 自然资源部, 国家林业和草原局.
Ministry of Natural Resources, National Forestry and Grassland Administration, 2020. Notice of the national forestry and grassland administration of the ministry of natural resources on the issuance of the special action plan for mangrove protection and restoration (2020-2025)[R]. Beijing: Ministry of Natural Resources, National Forestry and Grassland Administration (in Chinese).
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}