
西沙群岛银屿珊瑚礁的生态快速退化及其对鱼类的影响
|
雷明凤(1999—), 女, 广西桂林市人, 硕士研究生, 从事海洋生物生态研究。email: |
Copy editor: 林强
收稿日期: 2023-08-09
修回日期: 2023-10-22
网络出版日期: 2023-10-24
基金资助
国家自然科学基金项目(42090041)
国家自然科学基金项目(42030502)
The rapid ecological degradation and its impact on fish of the Yinyu Island in the Xisha Islands
Copy editor: LIN Qiang
Received date: 2023-08-09
Revised date: 2023-10-22
Online published: 2023-10-24
Supported by
National Natural Science Foundation of China(42090041)
National Natural Science Foundation of China(42030502)
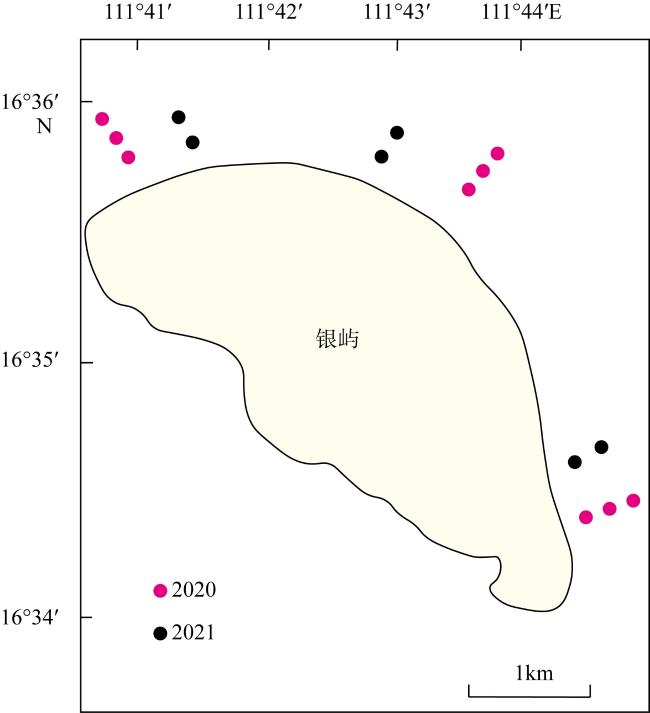
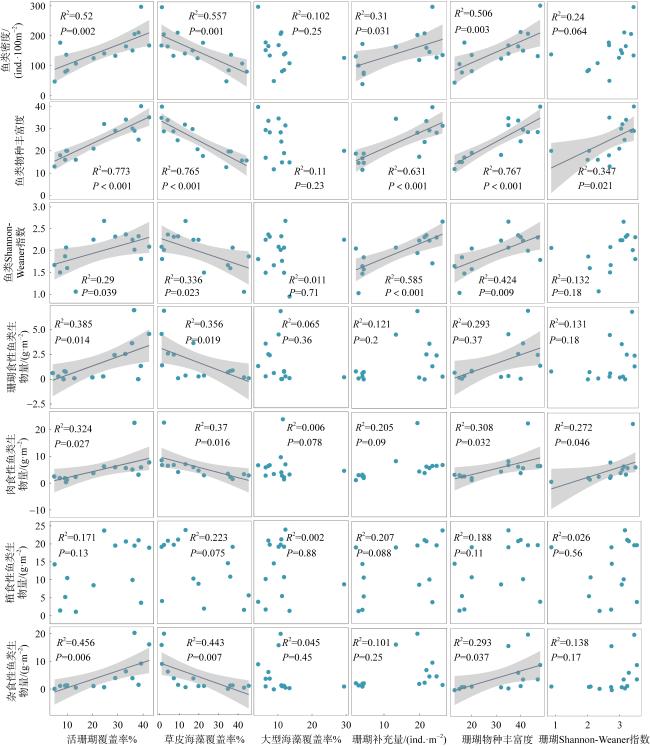
珊瑚礁生态系统退化能够改变珊瑚礁三维结构的复杂性及底栖生物组成, 从而对珊瑚礁鱼类群落产生重要影响。南海珊瑚礁在近几十年来处于快速退化之中, 但其退化对珊瑚礁鱼类群落造成的生态影响仍不清楚。西沙群岛珊瑚礁近年受高温和长棘海星暴发等的影响而出现生态退化, 因此, 本文以西沙群岛银屿珊瑚礁为研究对象, 通过水下录像样带调查和差异性检验、线性回归评估其珊瑚礁生态退化状况及其对珊瑚礁鱼类群落的影响。结果表明: 2020至2021年, 银屿活珊瑚覆盖率从33.27%下降到9.07%, 草皮海藻覆盖率从8.38%上升到36.50%, 珊瑚补充量从21.42ind.·m-2下降到3.64ind.·m-2, 珊瑚物种丰富度从40.00下降到16.83, 珊瑚Shannon-Weaner指数从3.26下降到2.14。鱼类密度从177.16ind·100m-2下降到105.07ind.·100m-2, 鱼类物种丰富度从30.56下降到17.17, 鱼类Shannon-Weaner指数从2.24下降到1.63, 珊瑚食性鱼类生物量从2.45g·m-2减少到0.43g·m-2, 肉食性鱼类生物量从7.35g·m-2减少到1.70g·m-2, 杂食性鱼类生物量从7.04g·m-2减少到0.95g·m-2, 鱼类总生物量从32.99g·m-2减少到11.66g·m-2。短短2年内, 银屿珊瑚礁已经被草皮海藻和大型海藻占据优势, 但是鱼类群落对珊瑚礁区的藻类并没有表现出自上而下的控制效应。在4种类群的鱼类中, 植食性鱼类受到生态退化的影响最小。珊瑚礁鱼类的物种丰富度和多样性均受限于珊瑚物种丰富度。基于以上结果, 推测银屿珊瑚礁的快速退化是导致珊瑚礁区鱼类群落物种多样性和密度下降, 改变珊瑚礁鱼类群落的结构的主要原因。希望未来进一步增加调查的频率和范围, 以便更深入地揭示鱼类群落与珊瑚礁生态变化的动态关系。
雷明凤 , 余克服 , 廖芝衡 , 陈飚 , 黄学勇 , 陈小燕 . 西沙群岛银屿珊瑚礁的生态快速退化及其对鱼类的影响[J]. 热带海洋学报, 2024 , 43(3) : 87 -99 . DOI: 10.11978/2023115
The degradation of coral reef ecosystem can alter the topographic complexity and biotic composition, thereby affecting the coral reef fish community significantly. Coral reefs in the South China Sea have undergone rapid degradation in recent decades, but the ecological consequences for coral reef fish community remain unclear. The coral reefs of the Xisha Islands have experienced ecological degradation in recent years due to the impacts of high temperature and the outbreak of crown-of-thorns starfish (CoTs). We conducted an ecological investigation on the Yinyu Island of the Xisha Islands, through underwater video transect survey, difference tests and liner regression analysis to assess the ecological degradation of the coral reef and its effect on the coral reef fish community. The results show that: from 2020 to 2021, the live coral cover on the Yinyu Island declined from 33.27%to 9.07%, while the turf algae cover increased from 8.38%to 36.50%. The coral recruitment, species richness and Shannon-Weaner index also decreased from 21.42 ind·m-2 to 3.64 ind·m-2, 40.00 to 16.83 and 3.26 to 2.14, respectively. The fish density, species richness, Shannon-Weaner index, declined from 1.77 ind.·m-2 to 1.05 ind.·m-2, 30.56 to 17.17 and 2.24 to 1.63, respectively. The biomass of corallivore, carnivore, omnivore and total biomass also declined from 2.45 g·m-2 to 0.43 g·m-2, 7.35 g·m-2 to 1.70 g·m-2, 7.04 g·m-2 to 0.95 g·m-2, and 32.99 g·m-2 to 11.66 g·m-2, respectively. Within two years, turf algae and macroalgae have dominated the Yinyu Island, but the fish community has not demonstrated top-down control of algae. Among the four functional groups in the whole fish community, the herbivore functional group is the least affected by ecological degradation. The coral species richness constrains the fish species richness and diversity. Based on the results above, we speculate that the rapid degradation of the Yinyu Island is the main reason for the decreased in species diversity, density and the change in the structure of coral reef fish communities. We hope to further increase the frequency and scope of investigations in the future, in order to reveal more deep dynamic relationship between fish communities and coral reef ecological changes.
Key words: coral reef; ecological degradation; fish community; Yinyu; Xisha Islands
图2 银屿珊瑚礁生态状况示意图a. 2020年生态状况: 活珊瑚覆盖率超过30%, 以块状的刺星珊瑚、滨珊瑚、蔷薇珊瑚和叶状的刺孔珊瑚以及枝状的杯形珊瑚为主; b. 2021年生态状况: 活珊瑚覆盖率低于10%, 以块状的滨珊瑚、盔形珊瑚和刺星珊瑚为主, 草皮海藻发育; c. 2021年隐藏在礁石内的长棘海星 Fig. 2 Ecological status of the Yinyu Island. (a) Ecological status in 2020: the live coral cover exceeded 30%, mainly consist of branching Pocillopora, massive Cyphastrea, Porites, Montipora and encrusting Echinpora; (b) ecological status in 2021: the live coral cover is less than 10%, mainly consist of massive Porites, Galaxea and Cyphastrea, with the development of turf algae; (c) crown-of-thorns starfish (CoTs) hidden within reefs in 2021 |
表1 2020和2021年银屿生态指标的平均值±标准误(SE)与差异性检验结果Tab. 1 Average of ecological indicators among the Yinyu Island over 2020 and 2021 and the results of difference test |
| 指标 | 2020年结果 | 2021年结果 | t | P |
|---|---|---|---|---|
| 活珊瑚覆盖率/% | 33.27±2.41 | 9.07±1.15 | 9.05 | <0.001 |
| 壳状珊瑚藻覆盖率/% | 29.67±2.79 | 24.97±4.32 | 0.92 | 0.384 |
| 草皮海藻覆盖率/% | 8.38±2.34 | 36.50±3.26 | -7.01 | <0.001 |
| 大型海藻覆盖率/% | 11.00±2.46 | 10.57±1.03 | 0.16 | 0.874 |
| 其他无脊椎动物覆盖率/% | 5.64±1.10 | 9.93±1.28 | -2.55 | 0.027 |
| 软珊瑚覆盖率/% | 5.36±1.06 | 1.07±0.10 | 52.00* | 0.004 |
| 礁石覆盖率/% | 3.04±0.53 | 1.23±0.40 | 2.72 | 0.018 |
| 碎石覆盖率/% | 0.84±0.26 | 0.70±0.47 | 0.27 | 0.795 |
| 沙覆盖率/% | 2.20±0.61 | 4.23±1.04 | -1.69 | 0.127 |
| 近期死亡珊瑚覆盖率/% | 0.60±0.25 | 1.73±0.47 | -2.13 | 0.066 |
| 珊瑚补充量/(ind.·m-2) | 21.42±1.25 | 3.64±0.36 | 13.6 | <0.001 |
| 珊瑚物种丰富度 | 40.00±1.82 | 16.83±1.14 | 10.8 | <0.001 |
| 珊瑚Shannon-Weaner指数 | 3.26±0.08 | 2.14±0.28 | 3.70 | 0.010 |
注: *软珊瑚覆盖率不满足方差齐性条件, 使用Wilcoxon检验, 其他指标均使用t检验, 粗体表示差异显著 |
表2 2020和2021年银屿珊瑚礁鱼类科、属、种及功能类群Tab. 2 Family, genus, species, and functional groups of fish on the Yinyu Island in 2020 and 2021 |
| 科 | 属 | 物种 | 功能类群 | 2020 | 2021 | |
|---|---|---|---|---|---|---|
| 刺盖鱼科 | 阿波鱼属 | 三点阿波鱼 | Apolemichthys trimaculatus | 肉食性 | + | + |
| 刺尻鱼属 | 海氏刺尻鱼 | Centropyge heraldi | 珊瑚食性 | + | ||
| 珠点刺尻鱼 | Centropyge vroliki | 珊瑚食性 | + | + | ||
| 刺尾鱼科 | 鼻鱼属 | 颊吻鼻鱼 | Naso lituratus | 植食性 | + | |
| 刺尾鱼属 | 橙斑刺尾鱼 | Acanthurus olivaceus | 植食性 | + | ||
| 横带刺尾鱼 | Acanthurus triostegus | 植食性 | + | |||
| 黄翼刺尾鱼 | Acanthurus xanthopterus | 植食性 | + | |||
| 日本刺尾鱼 | Acanthurus japonicus | 植食性 | + | |||
| 双斑刺尾鱼 | Acanthurus nigrofuscus | 植食性 | + | |||
| 栉齿刺尾鱼属 | 塞氏栉齿刺尾鱼 | Ctenochaetus cyanocheilus | 植食性 | + | + | |
| 双斑栉齿刺尾鱼 | Ctenochaetus binotatus | 植食性 | + | |||
| 栉齿刺尾鱼 | Ctenochaetus striatus | 植食性 | + | + | ||
| 单角鲀科 | 前孔鲀属 | 棘尾前孔鲀 | Cantherhines dumerilii | 肉食性 | + | |
| 仿石鲈科 | 胡椒鲷属 | 条斑胡椒鲷 | Plectorhinchus vittatus | 肉食性 | + | |
| 海鳝科 | 裸胸鳝属 | 斑点裸胸鳝 | Gymnothorax meleagris | 肉食性 | + | |
| 蝴蝶鱼科 | 蝴蝶鱼属 | 鞍斑蝴蝶鱼 | Chaetodon ulietensis | 杂食性 | + | |
| 斑带蝴蝶鱼 | Chaetodon punctatofasciatus | 珊瑚食性 | + | |||
| 叉纹蝴蝶鱼 | Chaetodon auripes | 杂食性 | + | |||
| 黑背蝴蝶鱼 | Chaetodon melannotus | 珊瑚食性 | + | |||
| 华丽蝴蝶鱼 | Chaetodon ornatissimus | 珊瑚食性 | + | |||
| 黄蝴蝶鱼 | Chaetodon xanthurus | 杂食性 | + | |||
| 密点蝴蝶鱼 | Chaetodon citrinellus | 珊瑚食性 | + | |||
| 三带蝴蝶鱼 | Chaetodon trifasciatus | 珊瑚食性 | + | + | ||
| 三纹蝴蝶鱼 | Chaetodon trifascialis | 珊瑚食性 | + | |||
| 蝴蝶鱼科 | 蝴蝶鱼属 | 项斑蝴蝶鱼 | Chaetodon adiergastos | 珊瑚食性 | + | |
| 珠蝴蝶鱼 | Chaetodon kleinii | 珊瑚食性 | + | |||
| 马夫鱼属 | 金口马夫鱼 | Heniochus chrysostomus | 珊瑚食性 | + | ||
| 四带马夫鱼 | Heniochus singularius | 珊瑚食性 | + | |||
| 镊口鱼属 | 黄镊口鱼 | Forcipiger flavissimus | 肉食性 | + | ||
| 霞蝶鱼属 | 多鳞霞蝶鱼 | Hemitaurichthys polylepis | 杂食性 | + | ||
| 金鳞鱼科 | 棘鳞鱼属 | 尾斑棘鳞鱼 | Sargocentron caudimaculatum | 肉食性 | + | |
| 锯鳞鱼属 | 孔锯鳞鱼 | Myripristis kuntee | 杂食性 | + | ||
| 金线鱼科 | 眶棘鲈属 | 双线眶棘鲈 | Scolopsis bilineata | 肉食性 | + | + |
| 镰鱼科 | 镰鱼属 | 镰鱼 | Zanclus cornutus | 肉食性 | + | |
| 鳞鲀科 | 凹纹鳞鲀属 | 黄边凹纹鳞鲀 | Xanthichthys auromarginatus | 杂食性 | + | |
| 多棘鳞鲀属 | 黄鳍多棘鳞鲀 | Sufflamen chrysopterum | 肉食性 | + | ||
| 角鳞鲀属 | 黑边角鳞鲀 | Melichthys vidua | 杂食性 | + | + | |
| 隆头鱼科 | 唇鱼属 | 横带唇鱼 | Cheilinus fasciatus | 肉食性 | + | + |
| 三叶唇鱼 | Cheilinus trilobatus | 肉食性 | + | |||
| 粗唇鱼属 | 横带粗唇鱼 | Hemigymnus fasciatus | 肉食性 | + | ||
| 大咽齿鱼属 | 珠斑大咽齿鱼 | Macropharyngodon meleagris | 肉食性 | + | ||
| 海猪鱼属 | 绿鳍海猪鱼 | Halichoeres marginatus | 肉食性 | + | + | |
| 圃海海猪鱼 | Halichoeres hortulanus | 肉食性 | + | |||
| 三斑海猪鱼 | Halichoeres trimaculatus | 肉食性 | + | + | ||
| 双睛斑海猪鱼 | Halichoeres biocellatus | 肉食性 | + | |||
| 尖唇鱼属 | 单带尖唇鱼 | Oxycheilinus unifasciatus | 肉食性 | + | + | |
| 尖嘴鱼属 | 杂色尖嘴鱼 | Gomphosus varius | 肉食性 | + | + | |
| 锦鱼属 | 鞍斑锦鱼 | Thalassoma hardwicke | 肉食性 | + | + | |
| 钝头锦鱼 | Thalassoma amblycephalum | 杂食性 | + | + | ||
| 纵纹锦鱼 | Thalassoma quinquevittatum | 肉食性 | + | + | ||
| 盔鱼属 | 露珠盔鱼 | Coris gaimard | 肉食性 | + | ||
| 裂唇鱼属 | 裂唇鱼 | Labroides dimidiatus | 肉食性 | + | + | |
| 双色裂唇鱼 | Labroides bicolor | 肉食性 | + | |||
| 胸斑裂唇鱼 | Labroides pectoralis | 肉食性 | + | |||
| 普提鱼属 | 双带普提鱼 | Bodianus bilunulatus | 肉食性 | + | ||
| 腋斑普提鱼 | Bodianus axillaris | 肉食性 | + | |||
| 伸口鱼属 | 伸口鱼 | Epibulus insidiator | 肉食性 | + | ||
| 丝隆头鱼属 | 黑缘丝隆头鱼 | Cirrhilabrus melanomarginatus | 杂食性 | + | + | |
| 蓝侧丝隆头鱼 | Cirrhilabrus cyanopleura | 肉食性 | + | |||
| 细鳞盔鱼属 | 狭带细鳞盔鱼 | Hologymnosus doliatus | 肉食性 | + | ||
| 紫胸鱼属 | 黑星紫胸鱼 | Stethojulis bandanensis | 肉食性 | + | ||
| 裸颊鲷科 | 齿颌鲷属 | 金带齿颌鲷 | Gnathodentex aureolineatus | 肉食性 | + | |
| 单列齿鲷属 | 单列齿鲷 | Monotaxis grandoculis | 肉食性 | + | ||
| 梅鲷科 | 梅鲷属 | 蓝黄梅鲷 | Caesio teres | 杂食性 | + | |
| 鮨科 | 侧牙鲈属 | 侧牙鲈 | Variola louti | 肉食性 | + | |
| 九棘鲈属 | 斑点九棘鲈 | Cephalopholis argus | 肉食性 | + | ||
| 豹纹九棘鲈 | Cephalopholis leopardus | 肉食性 | + | |||
| 尾纹九棘鲈 | Cephalopholis urodeta | 肉食性 | + | + | ||
| 石斑鱼属 | 六角石斑鱼 | Epinephelus hexagonatus | 肉食性 | + | ||
| 吻斑石斑鱼 | Epinephelus spilotoceps | 肉食性 | + | |||
| 鳍塘鳢科 | 鳍塘鳢属 | 黑尾鳍塘鳢 | Ptereleotris evides | 杂食性 | + | + |
| 线塘鳢属 | 大口线塘鳢 | Nemateleotris magnifica | 杂食性 | + | + | |
| 雀鲷科 | 凹牙豆娘鱼属 | 金凹牙豆娘鱼 | Amblyglyphidodon aureus | 杂食性 | + | |
| 豆娘鱼属 | 五带豆娘鱼 | Abudefduf vaigiensis | 杂食性 | + | ||
| 雀鲷科 | 光鳃鱼属 | 凡氏光鳃鱼 | Chromis vanderbilti | 杂食性 | + | + |
| 黄尾光鳃鱼 | Chromis xanthura | 杂食性 | + | |||
| 蓝绿光鳃鱼 | Chromis viridis | 杂食性 | + | |||
| 雀鲷科 | 光鳃鱼属 | 双斑光鳃鱼 | Chromis margaritifer | 杂食性 | + | + |
| 条尾光鳃鱼 | Chromis ternatensis | 杂食性 | + | + | ||
| 韦氏光鳃鱼 | Chromis weberi | 杂食性 | + | |||
| 椒雀鲷属 | 黑带椒雀鲷 | Plectroglyphidodon dickii | 杂食性 | + | + | |
| 眼斑椒雀鲷 | Plectroglyphidodon lacrymatus | 杂食性 | + | + | ||
| 金翅雀鲷属 | 圆尾金翅雀鲷 | Chrysiptera cyanea | 杂食性 | + | ||
| 眶锯雀鲷属 | 胸斑眶锯雀鲷 | Stegastes fasciolatus | 植食性 | + | + | |
| 雀鲷属 | 班卡雀鲷 | Pomacentrus bankanensis | 杂食性 | + | + | |
| 菲律宾雀鲷 | Pomacentrus philippinus | 杂食性 | + | + | ||
| 孔雀雀鲷 | Pomacentrus pavo | 杂食性 | + | + | ||
| 霓虹雀鲷 | Pomacentrus coelestis | 杂食性 | + | + | ||
| 王子雀鲷 | Pomacentrus vaiuli | 杂食性 | + | |||
| 双锯鱼属 | 克氏双锯鱼 | Amphiprion clarkii | 杂食性 | + | ||
| 宅泥鱼属 | 三斑宅泥鱼 | Dascyllus trimaculatus | 杂食性 | + | + | |
| 网纹宅泥鱼 | Dascyllus reticulatus | 杂食性 | + | + | ||
| 鲀科 | 叉鼻鲀属 | 黑斑叉鼻鲀 | Arothron nigropunctatus | 杂食性 | + | |
| 鳚科 | 橫口鳚属 | 黑带橫口鳚 | Plagiotremus tapeinosoma | 肉食性 | + | |
| 异齿鳚属 | 巴氏异齿鳚 | Ecsenius bathi | 植食性 | + | ||
| 虾虎鱼科 | 矶塘鳢属 | 胸斑矶塘鳢 | Eviota prasites | 杂食性 | + | |
| 箱鲀科 | 箱鲀属 | 粒突箱鲀 | Ostracion cubicus | 杂食性 | + | |
| 羊鱼科 | 副鲱鲤属 | 多带副绯鲤 | Parupeneus multifasciatus | 肉食性 | + | + |
| 鹦嘴鱼科 | 绿鹦嘴鱼属 | 污色绿鹦嘴鱼 | Chlorurus sordidus | 植食性 | + | + |
| 鹦嘴鱼属 | 弧带鹦嘴鱼 | Scarus dimidiatus | 植食性 | + | + | |
| 黄鞍鹦嘴鱼 | Scarus oviceps | 植食性 | + | |||
| 绿唇鹦嘴鱼 | Scarus forsteni | 植食性 | + | |||
| 网条鹦嘴鱼 | Scarus frenatus | 植食性 | + | |||
| 䱵科 | 副䱵属 | 副䱵 | Paracirrhites arcatus | 肉食性 | + | + |
| 金䱵属 | 尖头金䱵 | Cirrhitichthys oxycephalus | 肉食性 | + | + | |
| 䱵属 | 䱵 | Cirrhitus pinnulatus | 肉食性 | + | ||
注: +表示有这种鱼的记载 |
表3 2020和2021年银屿鱼类群落指标的平均值±标准误(SE)与差异性检验结果Tab. 3 Average of fish community indicators among Yinyu Island over the 2020 and 2021 two years and the results of difference test |
| 指标 | 2020年结果 | 2021年结果 | t | P |
|---|---|---|---|---|
| 鱼类密度/(ind.·100m-2) | 177.16±17.96 | 105.07±18.81 | 2.77 | 0.017 |
| 鱼类物种丰富度 | 30.56±1.86 | 17.17±1.11 | 6.17 | <0.001 |
| 鱼类Shannon-Weaner指数 | 2.24±0.08 | 1.63±0.14 | 3.37 | 0.005 |
| 珊瑚食性鱼类生物量/(g·m-2) | 2.45±0.78 | 0.43±0.14 | 2.56 | 0.027 |
| 肉食性鱼类生物量/(g·m-2) | 7.35±1.95 | 1.70±0.32 | 2.85 | 0.020 |
| 植食性鱼类生物量/(g·m-2) | 16.16±2.32 | 8.58±2.96 | 2.02 | 0.070 |
| 杂食性鱼类生物量/(g·m-2) | 7.04±2.32 | 0.95±0.22 | 2.61 | 0.031 |
| 鱼类总生物量/(g·m-2) | 32.99±5.58 | 11.66±2.90 | 3.39 | 0.006 |
| 鱼类群落长度谱斜率 | -0.19±0.21 | -0.15±0.01 | -3.78 | 0.008 |
| 鱼类群落长度谱截距 | 6.10±0.09 | 4.86±0.22 | 54* | <0.001 |
注: *鱼类群落长度谱截距不满足方差齐性条件, 使用Wilcoxon检验, 其他指标均使用t检验, 粗体表示差异显著 |
| [1] |
董俊德, 张燕英, 王友绍, 等, 2006. 南海三亚湾及邻近海域海洋蓝藻类群组成、分布及演替特征[J]. 热带海洋学报, 25(3): 40-46.
|
| [2] |
黄晖, 尤丰, 练健生, 等, 2011. 西沙群岛海域造礁石珊瑚物种多样性与分布特点[J]. 生物多样性, 19(6): 710-715.
|
| [3] |
黄晖, 2018. 西沙群岛珊瑚礁生物图册[M]. 北京:科学出版社: 2-370.
|
| [4] |
李太武, 2013. 海洋生物学[M]. 北京: 海洋出版社:180.
|
| [5] |
李颖虹, 黄小平, 岳维忠, 等, 2004. 西沙永兴岛珊瑚礁与礁坪生物生态学研究[J]. 海洋与湖沼, 35(2): 176-182.
|
| [6] |
李元超, 陈石泉, 郑新庆, 等, 2018. 永兴岛及七连屿造礁石珊瑚近10年变化分析[J]. 海洋学报, 40(8): 97-109.
|
| [7] |
李元超, 吴钟解, 梁计林, 等, 2019. 近15年西沙群岛长棘海星暴发周期及暴发原因分析[J]. 科学通报, 64(33): 3478-3484.
|
| [8] |
李媛洁, 陈作志, 张俊, 等, 2020. 西沙群岛七连屿礁栖鱼类物种和分类多样性[J]. 中国水产科学, 27(7): 815-823.
|
| [9] |
廖芝衡, 余克服, 王英辉, 2016. 大型海藻在珊瑚礁退化过程中的作用[J]. 生态学报, 36(21): 6687-6695.
|
| [10] |
廖芝衡, 2021. 南海珊瑚群落和底栖海藻的空间分布特征及其生态影响[D]. 南宁: 广西大学:39-40.
|
| [11] |
刘胜, 林先智, 张黎, 等, 2021. 南沙群岛珊瑚礁鱼类生态图册[M]. 北京: 科学出版社: 2-288 (in Chinese).
|
| [12] |
邱书婷, 刘昕明, 陈彬, 等, 2022. 西沙群岛珊瑚礁鱼类多样性及分布格局[J]. 海洋环境科学, 41(3): 395-401.
|
| [13] |
施祺, 严宏强, 张会领, 等, 2011. 西沙群岛永兴岛礁坡石珊瑚覆盖率的空间变化[J]. 热带海洋学报, 30(2): 10-17.
|
| [14] |
王腾, 石娟, 于洋飞, 等, 2023a. 西沙珊瑚礁鱼类研究进展和保护建议[J]. 生态学杂志, 42(7): 1755-1763.
|
| [15] |
王腾, 刘永, 李纯厚, 等, 2023b. 永兴岛附近海域珊瑚礁鱼类群落结构特征[J]. 水生生物学报, 47(4): 674-683.
|
| [16] |
吴钟解, 王道儒, 涂志刚, 等, 2011. 西沙生态监控区造礁石珊瑚退化原因分析[J]. 海洋学报, 33(4): 140-146.
|
| [17] |
杨位迪, 胡俊彤, 林柏岸, 等, 2018. 西沙赵述岛海域珊瑚礁鱼类物种多样性[J]. 厦门大学学报(自然科学版), 57(6): 819-826.
|
| [18] |
余克服, 2012. 南海珊瑚礁及其对全新世环境变化的记录与响应[J]. 中国科学: 地球科学, 42(8): 1160-1172.
|
| [19] |
张振冬, 邵魁双, 杨正先, 等, 2018. 西沙珊瑚礁生态承载状况评价研究[J]. 海洋环境科学, 37(4): 487-492.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}